
Alveolát - Alveolate
| Alveolát |
|
|---|---|

|
|
| Ceratium furca | |
|
Vědecká klasifikace |
|
| Doména: | Eukaryota |
| (nezařazeno): | Diaforetiky |
| Clade : | CAR |
| Clade : | SAR |
| Infrakingdom: |
Alveolata Cavalier-Smith , 1991 |
| Phyla | |
| Synonyma | |
|
|
Tyto alveolata (ve smyslu „s dutinami“) jsou skupinou prvoků , považován za hlavní clade a superphylum uvnitř Eukarya , a jsou také nazývány Alveolata .
Charakteristika
Nejpozoruhodnější sdílenou charakteristikou je přítomnost kortikálních (vnějších oblastí) alveolů (váčků). Jedná se o zploštělé váčky (váčky) zabalené do souvislé vrstvy těsně pod membránou a podporující ji, typicky vytvářející pružný pellikul (tenká kůže). V dinoflagellátech často tvoří pancéřové desky. Alveoláty mají mitochondrie s trubkovitými krystaly (hřebeny) a jejich bičíky nebo řasinky mají odlišnou strukturu.
Téměř všechny sekvenované mitochondriální genomy ciliatů a apicomplexií jsou lineární. Mitochondrie téměř všechny nesou vlastní mtDNA, ale se značně zmenšenými velikostmi genomu. Výjimkou jsou Cryptosporidium, kterým zbývá pouze mitosom ; ciliati; a Janouškovec et al 2013 prokázali, že Acavomonas se rozcházel časně, a tak si zachoval nějakou genově kódující mtDNA. Mitochondriální genom Babesia microti je kruhový. O tomto druhu je nyní také známo, že nepatří ani do rodů Babesia, ani Theileria, a bude pro něj muset být vytvořen nový rod.
Klasifikace
Alveolata zahrnuje přibližně 9 hlavních a vedlejších skupin, které mají velmi různorodou formu a je známo, že jsou příbuzné různými ultrastrukturálními a genetickými podobnostmi:
- Ciliates - velmi časté prvoki s mnoha krátkými řasinkami uspořádanými v řadách a dvěma jádry
- Acavomonidia
- Colponemidia
- Dinoflagellates sl - většinou mořské bičíkovce, z nichž mnohé mají chloroplasty
- Perkinsozoa
- Chromerida - mořský kmen fotosyntetických prvoků
- Colpodellida
- Voromonadida
- Apicomplexa -parazitické a sekundární nefotosyntetické prvoky, které postrádají axonemální pohybové struktury kromě gamet
Acavomonidia a Colponemidia byly dříve seskupeny jako kolponemidy, taxon nyní rozdělený na základě ultrastrukturální analýzy. Acavomonidia jsou blíže ke skupině dinoflagellate/perkinsid než Colponemidia. Neformální termín „colponemids“, jak je v současné době zastoupen, zahrnuje dvě nesesterské skupiny v Alveolatě: Acavomonidia a Colponemidia.
Apicomplexa a dinoflagelláty mohou být ve vzájemném bližším vztahu než s nálevníky. Oba mají plastidy a většina sdílí svazek nebo kužel mikrotubulů v horní části buňky. V apicomplexans to tvoří součást komplexu používaného pro vstup do hostitelských buněk, zatímco v některých bezbarvých dinoflagellátech tvoří stopku používanou k požití kořisti. S těmito dvěma skupinami úzce souvisí různé další rody, většinou bičíkovce s podobnou apikální strukturou. Patří sem volně žijící členové v Oxyrrhis a Colponema a paraziti v Perkinsus , Parvilucifera , Rastrimonas a ellobiopsids . V roce 2001 přímá amplifikace rRNA genu ve vzorcích mořského pikoplanktonu odhalila přítomnost dvou nových alveolátových linií, nazývaných skupina I a II. Skupina I nemá žádné kultivované příbuzné, zatímco skupina II je příbuzná dinoflagelátovému parazitu Amoebophrya , který byl dosud zařazen do řádu Syndiniales dinoflagellate.
Vztahy mezi některými z těchto hlavních skupin byly navrženy v 80. letech minulého století a konkrétní vztah mezi všemi třemi byl počátkem 90. let potvrzen genetickými studiemi, zejména Gajadharem a spol . Cavalier-Smith představil formální název Alveolata v roce 1991, ačkoli v té době ve skutečnosti považoval seskupení za paraphyletic asambláž, spíše než za monofyletickou skupinu.
Některé studie naznačovaly, že sem mohou patřit haplosporidy , většinou paraziti mořských bezobratlých, ale postrádají plicní sklípky a jsou nyní umístěny mezi Cercozoa .
Fylogeneze
Na základě kompilace následujících děl.
| Alveolata |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Nedávný výzkum naznačuje, že Dinoflagelllata se skutečně objevila v Perkinsea. Kromě toho se Perkinsea objevila v Apicomplexa.
Taxonomie
Alveolata Cavalier-Smith 1991 [Alveolatobiontes]
- Phylum Ciliophora Doflein 1901 stat. n. Copeland 1956 [ Ciliata Perty 1852 ; Infusoria Bütschli 1887 ; Ciliae, Ciliozoa, Cytoidea, Eozoa, Heterocaryota, Heterokaryota]
- Podkmen Postciliodesmatophora Gerassimova & Seravin 1976
- Třída Heterotrichea Stein 1859
- Třída Karyorelictea Corliss 1974
- Podkmen Intramacronucleata Lynn 1996
- Třída? Mesodiniea Chen a kol. 2015
- Infraphylum Lamellicorticata
- Třída Litostomatea Small & Lynn 1981
- Třída Armophorea Lynn 2004
- Třída Cariacotrichea Orsi a kol. 2011
- Třída Spirotrichea Bütschli 1889
- Infraphylum Ventrata Cavalier-Smith 2004 [Conthreep Lynn 2012 ]
- Objednat ? Discotrichida Chen et al. 2015
- Třída Protocruziea Chen et al. 2015 [Protocruziidia de Puytorac, Grain & Mignot 1987 ]
- Třída Colpodea Small & Lynn 1981
- Třída Nassophorea Small & Lynn 1981
- Třída Phyllopharyngea de Puytorac et al. 1974
- Třída Prostomatea Schewiakoff 1896
- Třída Plagiopylea Small & Lynn 1985 sensu Lynn 2008
- Třída Oligohymenophorea de Puytorac et al. 1974
- Podkmen Postciliodesmatophora Gerassimova & Seravin 1976
- Phylum Miozoa Cavalier-Smith 1987
- Subphylum Colponemidia Tikhonenkov, Mylnikov & Keeling 2013
- Třída Colponemea Cavalier-Smith 1993
- Subphylum Acavomonadia Tikhonenkov et al. 2014
- Třída Acavomonadea Tikhonenkov et al. 2014
- Podkmen Myzozoa Cavalier-Smith 2004
- Infraphylum Apicomplexa Levine 1970 emend. Adl a kol. 2005
- Objednat ? Vitrellida Cavalier-Smith 2017
- Třída? Myzomonadea Cavalier-Smith & Chao 2004 sensu Ruggiero et al. 2015
- Třída Chromerea
- Objednat Colpodellida Patterson & Zölffel 1991 [Spiromonadida Krylov & Mylnikov 1986 ]
- Nadtřídou Sporozoa Leuckart 1879 stat. listopad. Cavalier-Smith 2013 [Gamontozoa]
- Třída Blastogregarinida Chatton & Villeneuve 1936 [Blastogregarinina; Blastogregarinorina Chatton & Villeneuve 1936 ]
- Třída Paragregarea Cavalier-Smith 2014
- Třída Gregarinomorphea Grassé 1953
- Třída Coccidiomorphea Doflein 1901
- Infraphylum Dinozoa Cavalier-Smith 1981 emend. 2003
- Objednat ? Acrocoelida Cavalier-Smith & Chao 2004
- Objednat ? Rastromonadida Cavalier-Smith & Chao 2004
- Třída Squirmidea Norén 1999 stat. listopad. Cavalier-Smith 2014
- Superclass Perkinsozoa Norén et al. 1999 ss
- Třída Perkinsea Levine 1978 [Perkinsasida Levine 1978 ]
- Superclass Dinoflagellata Butschli 1885 stat. listopad. Cavalier-Smith 1999 sensu Cavalier-Smith 2013 [Dinozoa Cavalier-Smith 1981 ]
- Třída Pronoctilucea
- Třída Ellobiopsea Cavalier-Smith 1993 [Ellobiophyceae Loeblich III 1970 ; Ellobiopsida Whisler 1990 ]
- Třída Myzodinea Cavalier-Smith 2017
- Třída Oxyrrhea Cavalier-Smith 1987
- Třída Syndinea Chatton 1920 sl [Syndiniophyceae Loeblich III 1970 ss ; Syndina Cavalier-Smith ]
- Třída Endodinea Cavalier-Smith 2017
- Třída Noctiluciphyceae Fensome et al. 1993 [Noctilucae Haeckel 1866 ; Noctilucea Haeckel 1866 stat. listopad. ; Cystoflagellata Haeckel 1873 stat. listopad. Butschli 1887 ]
- Třída Dinophyceae Pascher 1914 [Peridinea Ehrenberg 1830 stat. listopad. Wettstein ]
- Infraphylum Apicomplexa Levine 1970 emend. Adl a kol. 2005
- Subphylum Colponemidia Tikhonenkov, Mylnikov & Keeling 2013
Rozvoj
Vývoj plastidů mezi alveoláty je zajímavý. Cavalier-Smith navrhl alveoláty vyvinuté z předchůdce obsahujícího chloroplasty, což také dalo podnět k Chromista ( hypotéza chromalveolátu ). Jiní vědci spekulovali, že v alveolátech původně chyběly plastidy a možná je dinoflageláty a Apicomplexa získaly samostatně. Nyní se však zdá, že alveoláty, dinoflageláty, Chromerida a heterokontní řasy získaly své plastidy z červené řasy s důkazem společného původu této organely ve všech těchto čtyřech kladech.
Vývoj
Bayesovský odhad předpokládá vývoj alveolátové skupiny před ~ 850 miliony let . Alveolata se skládá z Myzozoa , Ciliates a Colponemids. Jinými slovy, termín Myzozoa , což znamená „ odsávat obsah z kořisti“, lze neformálně aplikovat na společného předka podskupiny alveolátů, které nejsou ani ciliati, ani colponemidy. Predace na řasách je důležitým hybatelem evoluce alveolátů, protože může poskytnout zdroje pro endosymbiózu nových plastidů. Termín Myzozoa je proto praktický koncept pro sledování historie alveolátového kmene.
Předchůdci alveolátové skupiny mohli být fotosyntetičtí. Rodový alveolát pravděpodobně měl plastid . Chromeridy, apicomplexany a peridinin dinoflagelláty si tuto organelu zachovaly . Ještě o krok zpět, chromeridy, peridininové dinoflageláty a heterokontní řasy mají společnou monofyletickou plastidovou linii, tj. Získávají své plastidy z červené řasy , a tak se zdá pravděpodobné, že společným předkem alveolátů a heterokontů byla také fotosyntetika .
V jednom myšlenkový společného předka z dinoflageláty , apicomplexans , Colpodella , Chromerida a Voromonas byl myzocytotic dravec se dvěma heterodynamic flagella , mikropórů , trichocysts , rhoptries , micronemes , polárním kruhem a stočený otevřený oboustranný kónický . Zatímco společný předek alveolátů mohl mít také některé z těchto charakteristik, bylo argumentováno, že myzocytóza nebyla jednou z těchto charakteristik, protože ciliati přijímají kořist jiným mechanismem.
Probíhající diskuse se týká počtu membrán obklopujících plastid přes apicomplexany a určité dinoflageláty a původu těchto membrán. Tento ultrastrukturální charakter lze použít ke seskupení organismů a pokud je postava společná, může to znamenat, že phyla měla společného fotosyntetického předka. Na základě toho, že apicomplexans mají plastid obklopený 4 membránami a že peridinin dinoflagelláty mají plastid obklopený 3 membránami, Petersen et al. nebyli schopni vyloučit, že sdílený plastid stramenopil-alveolát mohl být několikrát recyklován v alveolátovém kmeni, přičemž zdrojem jsou dárci stramenopile-alveolátu, prostřednictvím mechanismu požití a endosymbiózy .
Ciliate jsou modelové alveoláty, které byly geneticky studovány ve velké hloubce po nejdelší dobu jakékoli alveolátové linie. Mezi eukaryoty jsou neobvyklé v tom, že reprodukce zahrnuje mikronukleus a makronukleus . Jejich reprodukce se snadno studuje v laboratoři a historicky z nich udělala modelový eukaryot. Jejich vývoj jako kmen, který byl zcela dravý a postrádal jakýkoli zbytkový plastid, ukazuje, jak jsou predace a autotrofie v dynamické rovnováze a že se rovnováha může hýbat tak či onak v místě vzniku nového kmene od mixotrofních předků, což způsobuje jednu schopnost být ztracen.
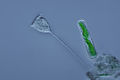
Vorticella ( Ciliophora ) (vlevo)

Plasmodium falciparum ( Apicomplexa ) v krvi




