
Cochlea - Cochlea
| Kochlea | |
|---|---|
 Průřez hlemýždě
| |
|
Části vnitřního ucha, zobrazující hlemýžď
| |
| Podrobnosti | |
| Výslovnost | / K ɒ k . l i ə / |
| Část | Vnitřní ucho |
| Systém | Sluchový systém |
| Identifikátory | |
| latinský | Kochlea |
| Pletivo | D003051 |
| NeuroLex ID | birnlex_1190 |
| TA98 | A15.3.03.025 |
| TA2 | 6964 |
| FMA | 60201 |
| Anatomická terminologie | |

 |
| Tento článek je jedním ze série dokumentující anatomii |
| Lidské ucho |
|---|

Hlemýžď je součástí vnitřního ucha zapojených do slyšení . Jedná se o spirálovitou dutinu v kostnatém labyrintu , u člověka se kolem jeho osy, modiolus, dělá 2,75 otáčky . Základní složkou hlemýždě je Cortiho orgán, smyslový orgán sluchu, který je distribuován podél přepážky oddělující tekutinové komory ve stočené kuželovité trubičce hlemýždě.
Název kochlea pochází ze starořeckého κοχλίας (kokhlias) „spirála, skořápka šneka“.
Struktura

Cochlea (množné číslo je cochleae) je spirálovitá, dutá, kuželovitá komora kosti, ve které se vlny šíří od základny (poblíž středního ucha a oválného okénka ) k vrcholu (vrchol nebo střed spirály). Spirálovitý kanál hlemýždě je část kostnatého labyrintu vnitřního ucha, která je přibližně 30 mm dlouhá a dělá 2¾ otáčky kolem modiolu. Kochleární struktury zahrnují:
- Tři stupnice nebo komory:
- vestibulární potrubí nebo Scala vestibuli (obsahující perilymfa ), který leží nadřazený kochleární kanálu a dosedá na oválné okno
- bubínku potrubí nebo scala tympani (obsahující perilymfa), který leží nižší než kochleární potrubí a končí na kole okně
- na kochleární potrubí nebo Scala média (obsahující endolymph ) oblast koncentrace iontů draslíku vysoká, že stereocilie z vlasových buněk promítnout
- Helicotrema , místo, kde bubínku potrubí a vestibulární sloučení potrubí, na vrcholu hlemýždě
- Reissnerova membrána , která odděluje vestibulární kanál od kochleárního kanálu
- Kostní spirála lamina , hlavní konstrukční prvek, který odděluje kochleární kanál z ušního kanálku
- Bazilární membrány , hlavní konstrukční prvek, který odděluje kochleární kanál z ušního kanálku a určuje mechanické šíření vln vlastnosti kochleární přepážky
- Cortiho orgán , senzorická epitel, buněčná vrstva na bazilární membrány, ve které jsou smyslové vlasové buňky napájen potenciální rozdíl mezi perilymph a endolymph
- vláskové buňky , smyslové buňky v Cortiho orgánu, zakončené vlasovými strukturami zvanými stereocilie
- Spirála vaz .
Hlemýžď je část vnitřního ucha, která vypadá jako skořápka šneka (hlemýžď je řecký výraz pro šneka). Hlemýžď přijímá zvuk ve formě vibrací, které způsobují pohyb stereocilií. Stereocilie pak převádí tyto vibrace na nervové impulsy, které jsou přijímány do mozku k interpretaci. Dvě ze tří tekutých sekcí jsou kanály a třetí je „Cortiho orgán“, který detekuje tlakové impulsy, které cestují podél sluchového nervu do mozku. Tyto dva kanály se nazývají vestibulární kanál a tympanický kanál.
Mikroanatomie
Stěny duté hlemýždě jsou kostěné, s tenkou, jemnou výstelkou epiteliální tkáně . Tato stočená trubice je po většinu své délky rozdělena vnitřní membránovou přepážkou. Tato dělící se membrána tvoří dva vnější prostory naplněné tekutinou (kanály nebo šupiny ). V horní části šnekovitých spirálových trubek dochází ke změně směru tekutiny, čímž se vestibulární kanál změní na tympanický kanál. Tato oblast se nazývá helicotrema. Toto pokračování v helicotrema umožňuje, aby se tekutina tlačila do vestibulárního kanálu oválným oknem, aby se pohybovala zpět pohybem v bubínkovém kanálu a vychýlením kulatého okna; protože tekutina je téměř nestlačitelná a kostnaté stěny jsou tuhé, je nezbytné, aby konzervovaný objem tekutiny někam vycházel.
Podélná přepážka, která rozděluje většinu hlemýždě, je sama trubicí naplněnou tekutinou, třetím „kanálem“. Tento centrální sloup se nazývá kochleární potrubí. Jeho tekutina, endolymfa, také obsahuje elektrolyty a bílkoviny, ale je chemicky zcela odlišná od perilymfy. Zatímco perilymfa je bohatá na ionty sodíku, endolymfa je bohatá na ionty draslíku, které produkují iontový , elektrický potenciál.
Vláskové buňky jsou uspořádány ve čtyřech řadách v Cortiho orgánu po celé délce kochleární cívky. Tři řady se skládají z vnějších vlasových buněk (OHC) a jedna řada se skládá z vnitřních vláskových buněk (IHC). Vnitřní vláskové buňky poskytují hlavní nervový výstup hlemýždě. Vnější vláskové buňky místo toho převážně „přijímají“ neurální vstupy z mozku, což ovlivňuje jejich pohyblivost jako součást mechanického „předzesilovače“ kochley. Vstup do OHC je z olivového těla prostřednictvím mediálního olivocochlearního svazku.
Kochleární kanál je sám o sobě téměř stejně složitý jako samotné ucho. Kochleární kanál je ze tří stran ohraničen bazilární membránou , stria vascularis a Reissnerovou membránou. Rýha vascularis je bohatá postel kapilár a sekrečních buněk; Reissnerova membrána je tenká membrána, která odděluje endolymfu od perilymfy; a bazilární membrána je mechanicky poněkud tuhá membrána, podporující receptorový orgán pro sluch, Cortiho orgán, a určuje vlastnosti mechanického šíření vln kochleárního systému.
Funkce

Hlemýžď je naplněn vodnatou kapalinou, endolymfou , která se pohybuje v reakci na vibrace přicházející ze středního ucha oválným oknem. Jak se tekutina pohybuje, pohybuje se kochleární přepážka (bazilární membrána a Cortiho orgán); tisíce vlasových buněk cítí pohyb prostřednictvím svých stereocilií a převádějí tento pohyb na elektrické signály, které jsou prostřednictvím neurotransmiterů sdělovány mnoha tisícům nervových buněk. Tyto primární sluchové neurony transformují signály na elektrochemické impulsy známé jako akční potenciály , které putují podél sluchového nervu do struktur v mozkovém kmeni pro další zpracování.
Sluch
Třmínek (třmen) kůstek kost středního ucha přenáší vibrace k Fenestra ovalis (oválné okno) na vnější straně hlemýždě, který vibruje perilymph v vestibulárního kanálu (horní komoře hlemýždě). Ossicles jsou nezbytné pro účinné spojení zvukových vln do hlemýždě, protože prostředí hlemýždě je systém tekutina -membrána a k pohybu zvuku vlnami tekutina -membrána je zapotřebí většího tlaku než vzduchem. Zvýšení tlaku je dosaženo snížením poměru ploch od bubínku (bubnu) k oválnému okénku ( stapová kost) o 20. Protože tlak = síla/plocha, vede k nárůstu tlaku asi 20krát z původního tlaku zvukové vlny ve vzduchu. Tento zisk je formou přizpůsobení impedance - aby se zvuková vlna cestující vzduchem přizpůsobila té, která cestuje v systému tekutina -membrána.
Na základně hlemýždě končí každý „kanál“ membránovým portálem, který směřuje do dutiny středního ucha: Vestibulární kanál končí u oválného okénka , kde sedí podnožka stapů . Podnožka vibruje, když je tlak přenášen přes ossikulární řetěz. Vlna v perilymfě se pohybuje pryč od stupačky a směrem k helicotrema . Vzhledem k tomu, že tyto tekutinové vlny pohybují kochleární přepážkou, která odděluje kanály nahoru a dolů, mají vlny odpovídající symetrickou část v perilymfě tympanického kanálu, který končí u kulatého okna a vyboulí se ven, když se oválné okno vyboulí dovnitř.
Perilymfa ve vestibulárním kanálu a endolymfa v kochleárním potrubí působí mechanicky jako jediné potrubí, přičemž je odděluje pouze velmi tenká Reissnerova membrána . Vibrace endolymfy v kochleárním potrubí vytlačují bazilární membránu ve vzoru, který vrcholí vzdálenost od oválného okna v závislosti na frekvenci zvukové vlny. Cortiho orgán vibruje v důsledku vnějších vlasových buněk dále zesilovat tyto vibrace. Buňky vnitřního vlasu jsou poté vytlačeny vibracemi v tekutině a depolarizují se přílivem K+ prostřednictvím jejich kanálů spojených špičkou a vysílají své signály prostřednictvím neurotransmiteru do primárních sluchových neuronů spirálního ganglia .
Vláskové buňky v Cortiho orgánu jsou naladěny na určité zvukové frekvence podle jejich umístění v hlemýždi, vzhledem ke stupni tuhosti bazilární membrány. Tato tuhost je mimo jiné dána tloušťkou a šířkou bazilární membrány, která je po délce hlemýždě nejtuhší nejblíže jejímu začátku u oválného okénka, kde stapes zavádí vibrace vycházející z bubínku. Jelikož je jeho tuhost vysoká, umožňuje pohybem bazilární membrány, a tím i vláskových buněk, pouze vysokofrekvenční vibrace. Čím dále vlna postupuje směrem k vrcholu hlemýždě ( helicotrema ), tím méně je bazilární membrána tuhá; nižší frekvence se tedy pohybují po trubce a méně tuhá membrána se jimi pohybuje nejsnadněji tam, kde to snížená tuhost umožňuje: to znamená, že jak se bazilární membrána stává stále méně tuhou, vlny zpomalují a lépe reaguje na nižší frekvence. U savců je navíc cochlea stočená, což bylo prokázáno, že zvyšuje nízkofrekvenční vibrace, když procházejí cívkou naplněnou tekutinou. Toto prostorové uspořádání příjmu zvuku se označuje jako tonotopy .
U velmi nízkých frekvencí (pod 20 Hz) se vlny šíří po celé trase hlemýždě - různě po vestibulárním a tympanickém kanálu až po helicotrema . Takto nízké frekvence stále do určité míry aktivují Cortiho orgán, ale jsou příliš nízké na to, aby vyvolaly vnímání výšky . Vyšší frekvence se nešíří do helicotrema , kvůli tonotopii zprostředkované tuhostí.
Velmi silný pohyb bazilární membrány v důsledku velmi hlasitého hluku může způsobit odumírání vlasových buněk. To je častou příčinou částečné ztráty sluchu a je důvodem, proč uživatelé střelných zbraní nebo těžkých strojů často nosí chrániče sluchu nebo ucpávky do uší .
Amplifikace vlasových buněk
Hlemýžď nejen „přijímá“ zvuk, ale zdravý hlemýžď v případě potřeby zvuk generuje a zesiluje. Tam, kde organismus potřebuje mechanismus pro slyšení velmi slabých zvuků, se kochlea zesiluje reverzní transdukcí OHC a převádí elektrické signály zpět na mechanické v konfiguraci s pozitivní zpětnou vazbou. OHC mají na svých vnějších membránách proteinový motor zvaný prestin ; vytváří další pohyb, který se spojuje zpět s vlnou tekutina -membrána. Tento „aktivní zesilovač“ je nezbytný pro schopnost ucha zesílit slabé zvuky.
Aktivní zesilovač také vede k fenoménu zvukových vln vibrací emitovaných z hlemýždě zpět do zvukovodu středním uchem (otoakustické emise).
Otoakustické emise
Otoakustické emise jsou způsobeny vlnou opouštějící hlemýžď oválným oknem a šířící se zpět středním uchem do bubínku a ven ze zvukovodu, kde jej lze zachytit mikrofonem. Otoakustické emise jsou v některých typech testů na poruchy sluchu důležité , protože jsou přítomny, když kochlea funguje dobře, a méně, když trpí ztrátou aktivity OHC.
Role mezerových křižovatek
Proteiny s mezerou , nazývané konexiny , exprimované v hlemýždi, hrají důležitou roli ve sluchovém fungování. Bylo zjištěno, že mutace v genech spojujících mezery způsobují syndromickou a nesyndromickou hluchotu. Některé konexiny, včetně konexinu 30 a konexinu 26 , převládají ve dvou odlišných mezerových spojovacích systémech nacházejících se v hlemýždi. Síťová spojovací mezera mezi epiteliálními buňkami spojuje nesenzorické epiteliální buňky, zatímco spojovací síť s mezerovou spojovací tkání spojuje buňky pojivové tkáně. Mezerové kanály recyklují ionty draslíku zpět do endolymfy po mechanotransdukci ve vlasových buňkách . Důležité je, že mezi kochleárními podpůrnými buňkami jsou mezerové spojovací kanály, nikoli však sluchové vláskové buňky .
Klinický význam
Ztráta sluchu
Bionika
V roce 2009 inženýři z Massachusettského technologického institutu vytvořili elektronický čip, který dokáže rychle analyzovat velmi široký rozsah rádiových frekvencí a přitom využívat jen zlomek energie potřebné pro stávající technologie; jeho design specificky napodobuje hlemýžď.
Ostatní zvířata
Svinutá forma hlemýždě je jedinečná pro savce . U ptáků a jiných obratlovců , kteří nejsou savci , se oddělení obsahující smyslové buňky pro sluch občas nazývá také „kochlea“, přestože není stočeno. Místo toho tvoří slepou trubici, nazývanou také kochleární potrubí. Tento rozdíl se zjevně vyvíjel souběžně s rozdíly ve frekvenčním rozsahu sluchu mezi savci a savci, kteří nejsou savci. Vynikající frekvenční rozsah u savců je částečně způsoben jejich unikátním mechanismem pre-amplifikace zvuku aktivními vibracemi buněk a těl vnějších vláskových buněk . Rozlišení frekvence není u savců lepší než u většiny ještěrek a ptáků, ale horní hranice frekvence je - někdy mnohem - vyšší. Většina druhů ptáků neslyší nad 4–5 kHz, aktuálně známé maximum je u sovy pálené ~ 11 kHz. Někteří mořští savci slyší až 200 kHz. Dlouhý stočený oddíl, spíše než krátký a rovný, poskytuje více prostoru pro další oktávy sluchového dosahu a umožnil některé z vysoce odvozených chování zahrnujících savčí sluch.
Jelikož by studium hlemýždě mělo být zásadně zaměřeno na úrovni vláskových buněk, je důležité si všimnout anatomických a fyziologických rozdílů mezi vláskovými buňkami různých druhů. Například u ptáků jsou místo vnějších a vnitřních vláskových buněk vysoké a krátké vláskové buňky. Pokud jde o tato srovnávací data, existuje několik podobností. Za prvé, vysoká vlásková buňka má velmi podobnou funkci jako vnitřní vlásková buňka a krátká vlásková buňka postrádá aferentní inervaci vláken sluchového nervu, připomíná vnější vláskovou buňku. Jeden nevyhnutelné rozdílem však je, že i když všechny vlasové buňky jsou připojeny k tektoriální membrána u ptáků, tak vnější vlasové buňky jsou připojeny k tektoriální membrána u savců.
Dějiny
Název cochlea je odvozen z latinského slova pro skořápku šneka , což je zase z řeckého κοχλίας kokhlias („šnek, šroub“), z κόχλος kokhlos („spirálová skořápka“) v odkazu na jeho stočený tvar; kochlea je u savců svinuta s výjimkou monotremů .
Další obrázky

Pravý kostnatý labyrint. Boční pohled

Interiér pravého kostního labyrintu.
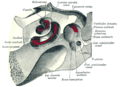
Hlemýžď a zádveří, při pohledu shora.
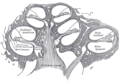
Průřez hlemýždě.




Viz také
- Kostnatý labyrint
- Membránový labyrint
- Kochleární implantát
- Kochleární nerv
- Kochleární jádra
- Evoluce hlemýždě
- Účinky hluku na zdraví
Reference
Další čtení
- Dallos, Peter; Popper, Arthur N .; Fay, Richard R. The Cochlea .
- Imbert, Michel; Kay, RH (1992). Konkurz . ISBN 9780262023313.
- Jahn, Anthony F .; Santos-Sacchi, Joseph (2001). Fyziologie ucha . ISBN 9781565939943.
- Roeser, Ross J .; Valente, Michael; Hosford-Dunn, Holly (2007). Audiologie . ISBN 9781588905420.
externí odkazy
- Cochlea v Americké národní knihovně lékařských lékařských oborových nadpisů (MeSH)
- „Promenade 'Round the Cochlea“ od R. Pujola, S. Blatrixe, T. Pujola a kol. na univerzitě v Montpellier
- „Histologická videa z ucha“