
Let hmyzem - Insect flight

Hmyz je jedinou skupinou bezobratlých, u kterých se vyvinula křídla a úlet . Hmyz poprvé letěl v karbonu , asi před 350 až 400 miliony let, což z něj činí první zvířata, která vyvinula let. Křídla se mohla vyvinout z přívěsků po stranách stávajících končetin, které již měly nervy, klouby a svaly používané k jiným účelům. Ty mohly být původně použity k plavbě po vodě nebo ke zpomalení klesání při klouzání.
Dvě skupiny hmyzu, vážky a jepice , mají letové svaly připevněné přímo na křídlech. U jiného okřídleného hmyzu se k hrudníku připevňují letové svaly, které jej oscilují, aby přiměly křídla porazit. Z tohoto hmyzu někteří ( mouchy a někteří brouci ) dosahují velmi vysokých frekvencí úderů křídel vývojem „asynchronního“ nervového systému, ve kterém hrudník osciluje rychleji než rychlost nervových impulzů.
Některý velmi malý hmyz nevyužívá aerodynamiku ustáleného stavu , ale mechanismus tleskání a mávání Weis-Fogh , který generuje velké vztlakové síly na úkor opotřebení křídel. Mnoho hmyzu se může vznášet, udržovat si výšku a kontrolovat svoji polohu. Některý hmyz, jako jsou můry, má přední křídla spojená se zadními křídly, takže mohou fungovat současně.
Mechanismy
Přímý let
Na rozdíl od jiných hmyzu, křídlové svaly Ephemeroptera (jepice) a Odonata (vážky a damselflies) vložit přímo na křídle bází, které jsou zavěšeny tak, že malý pohyb směrem dolů křídla základny zvedne křídlo sám nahoru, stejně jako veslování přes vzduch. Vážky a motýlice mají přední a zadní křídla podobného tvaru a velikosti. Každý pracuje samostatně, což poskytuje určitý stupeň jemné kontroly a pohyblivosti, pokud jde o prudkost, s jakou mohou měnit směr a rychlost, což u jiného létajícího hmyzu není vidět. Odonáti jsou letečtí predátoři a vždy lovili jiný vzdušný hmyz.

Přímý let: svaly připevněné k křídlům. Pouze velký hmyz


Nepřímý let
Kromě dvou řádů s přímými letovými svaly létá veškerý ostatní okřídlený hmyz pomocí jiného mechanismu, který zahrnuje svaly nepřímého letu. Tento mechanismus se vyvinul jednou a je určujícím znakem ( synapomorfie ) pro infratřídu Neoptera ; to odpovídá, pravděpodobně ne náhodou, vzhledu mechanismu skládání křídel, který umožňuje neopteranskému hmyzu sklopit křídla zpět přes břicho, když je v klidu (ačkoli tato schopnost byla v některých skupinách ztracena sekundárně, například u motýlů ) .
Co vše však Neoptera sdílí, je způsob, jakým svaly v hrudníku fungují: tyto svaly se místo přichycení ke křídlům připevňují k hrudníku a deformují jej; protože křídla jsou prodloužením hrudního exoskeletu , deformace hrudníku způsobí pohyb křídel také. Sada podélných svalů podél zad stlačuje hrudník zepředu dozadu, což způsobí, že hřbetní povrch hrudníku ( notum ) se skloní vzhůru a křídla se převrátí dolů. Další sada svalů od tergumu k hrudní kosti táhne notum opět dolů, což způsobí převrácení křídel nahoru.
Nepřímý let: svaly způsobují, že hrudník u většiny hmyzu osciluje


Hmyz, který porazil křídla méně než stokrát za sekundu, používá synchronní sval. Synchronní sval je typ svalu, který se smršťuje jednou pro každý nervový impuls. To obecně produkuje méně energie a je méně účinné než asynchronní sval, což odpovídá nezávislému vývoji asynchronních letových svalů v několika oddělených kladích hmyzu.
Hmyz, který rychleji mlátí křídly, jako čmelák , používá asynchronní sval; toto je typ svalu, který se smršťuje více než jednou na nervový impuls. Toho je dosaženo tím, že sval je znovu stimulován ke kontrakci uvolněním napětí ve svalu, což může nastat rychleji než pouhou jednoduchou nervovou stimulací samotnou. To umožňuje frekvenci úderů křídla překročit rychlost, s jakou může nervový systém vysílat impulsy. Asynchronní sval je jedním z konečných zdokonalení, které se objevilo u některých vyšších Neoptera ( Coleoptera , Diptera a Hymenoptera ). Celkový efekt je ten, že mnoho vyšších Neopter dokáže porazit křídla mnohem rychleji než hmyz s přímými letovými svaly.
Aerodynamika
Existují dva základní aerodynamické modely letu hmyzu: vytvoření víru na náběžné hraně a použití tleskání a házení.
Vortex náběžné hrany
Většina hmyzu používá metodu, která vytváří spirálovitý vír na náběžné hraně . Tato mávající křídla se pohybují dvěma základními polovičními tahy. Downstroke začíná nahoru a zpět a je ponořen dolů a dopředu. Poté se křídlo rychle převrátí ( supinace ) tak, aby přední hrana směřovala dozadu. Tah nahoru pak tlačí křídlo nahoru a dozadu. Poté se křídlo znovu převrátí ( pronace ) a může dojít k dalšímu stáhnutí. Frekvenční rozsah v hmyzu se synchronními letových svalů je typicky od 5 do 200 hertzů (Hz). U osob s asynchronními letovými svaly může frekvence úderu křídla přesáhnout 1 000 Hz. Když se hmyz vznáší, oba tahy trvají stejně dlouho. Pomalejší sestup však poskytuje tah .
Identifikace hlavních sil je zásadní pro pochopení letu hmyzu. První pokusy o pochopení mávání křídly předpokládaly kvazi-ustálený stav. To znamená, že průtok vzduchu přes křídlo v daném okamžiku byl považován za stejný, jako by byl proud přes neklapající, ustálené křídlo ve stejném úhlu náběhu. Rozdělením mávajícího křídla do velkého počtu nehybných poloh a následnou analýzou každé polohy by bylo možné vytvořit časovou osu okamžitých sil na křídle v každém okamžiku. Vypočítaný vztlak byl trojnásobně příliš malý, takže vědci si uvědomili, že musí existovat nestabilní jevy poskytující aerodynamické síly. Existuje několik vyvíjejících se analytických modelů, které se pokoušejí přiblížit tok v blízkosti mávajícího křídla. Někteří vědci předpovídali vrcholy sil při supinaci. S dynamicky zmenšeným modelem ovocné mušky byly tyto předpovězené síly později potvrzeny. Jiní tvrdili, že vrcholy sil během supinace a pronace jsou způsobeny neznámým rotačním efektem, který se zásadně liší od translačních jevů. S tímto argumentem existuje určitý nesouhlas. Prostřednictvím výpočetní dynamiky tekutin někteří vědci tvrdí, že neexistuje žádný rotační efekt. Tvrdí, že vysoké síly jsou způsobeny interakcí s bděním prolitým předchozím úderem.
Podobně jako u výše zmíněného rotačního efektu nejsou jevy spojené s mávajícími křídly zcela pochopeny nebo dohodnuty. Protože každý model je přibližný, různé modely vynechávají efekty, které jsou považovány za zanedbatelné. Například Wagnerův efekt , jak navrhl Herbert A. Wagner v roce 1925, říká, že cirkulace pomalu stoupá do ustáleného stavu kvůli viskozitě, když je nakloněné křídlo zrychleno z klidu. Tento jev by vysvětlil hodnotu zdvihu, která je menší, než se předpokládá. Typickým případem bylo nalezení zdrojů pro přidaný výtah. Bylo argumentováno, že tento účinek je zanedbatelný pro tok s Reynoldsovým číslem, které je typické pro létání hmyzu. Reynoldsovo číslo je mírou turbulencí ; tok je laminární (plynulý), když je Reynoldsovo číslo nízké, a turbulentní, když je vysoký. Minimálně v jednom modelu byl Wagnerův efekt vědomě ignorován. Jedním z nejdůležitějších jevů, ke kterým dochází během letu hmyzu, je sání náběžné hrany. Tato síla je významná pro výpočet účinnosti. DG Ellis a JL Stollery v roce 1988 poprvé navrhli koncept sání náběžné hrany, aby popsali vortexový vztlak na ostrohranných delta křídlech . Při vysokých úhlech útoku se tok odděluje přes náběžnou hranu, ale znovu se připojuje, než dosáhne odtokové hrany. V této bublině odděleného toku je vír. Protože je úhel náběhu tak vysoký, hodně hybnosti se přenáší dolů do toku. Tyto dvě funkce vytvářejí velké množství vztlakové síly a také dodatečný odpor. Důležitou vlastností je však výtah. Vzhledem k tomu, že se tok oddělil, přesto stále poskytuje velké množství vztlaku, tento jev se nazývá zpoždění při zastavení , kterého si poprvé všiml na vrtulích letadel H. Himmelskamp v roce 1945. Tento účinek byl pozorován při mávání létajícím hmyzem a bylo prokázáno, že je schopen zajištění dostatečného zdvihu, který by zohlednil nedostatek v kvazi-ustálených modelech. Tento efekt využívají kanoisté při tahu na lezení .
Všechny efekty na mávající křídlo lze omezit na tři hlavní zdroje aerodynamických jevů: vír náběžné hrany, aerodynamické síly v ustáleném stavu na křídlo a kontakt křídla s jeho probuzením z předchozích úderů. Velikost létajícího hmyzu se pohybuje od asi 20 mikrogramů do asi 3 gramů. Jak se zvyšuje tělesná hmotnost hmyzu, zvětšuje se plocha křídla a snižuje se frekvence úderů křídel. U většího hmyzu může být Reynoldsovo číslo (Re) až 10 000, kde proudění začíná být turbulentní. U menšího hmyzu to může být až 10. To znamená, že viskózní efekty jsou pro menší hmyz mnohem důležitější.


Další zajímavou vlastností letu hmyzu je náklon těla. Jak se rychlost letu zvyšuje, tělo hmyzu má sklon naklonit nos dolů a stát se více horizontální. Tím se zmenší čelní plocha a tím i odpor těla. Vzhledem k tomu, že se vzrůstající rychlostí vpřed také zvyšuje odpor, hmyz zefektivňuje svůj let, protože tato účinnost je stále potřebnější. Kromě toho, díky změně geometrického úhlu nárazu na sestřel, je hmyz schopen udržet svůj let s optimální účinností při co největším počtu manévrů. Vývoj celkového tahu je relativně malý ve srovnání se vztlakovými silami. Zvedací síly mohou být více než trojnásobkem hmotnosti hmyzu, zatímco tah i při nejvyšších rychlostech může být až 20% hmotnosti. Tato síla se vyvíjí především prostřednictvím méně silného vztlaku mávajícího pohybu.
Tleskat a mrskat


Clap and fling, neboli mechanismus Weis-Fogh, objevený dánským zoologem Torkelem Weis-Foghem , je metoda generování výtahu využívaná při letu malým hmyzem. Jak se velikosti hmyzu stávají menší než 1 mm, viskózní síly se stávají dominantní a účinnost generování výtahu z profilu křídla se drasticky snižuje. Počínaje polohou tleskání se obě křídla od sebe odtrhnou a otáčejí se kolem odtokové hrany. Křídla se poté oddělí a vodorovně se smetou až do konce dolů. Dále křídla pronikají a využívají náběžnou hranu během pohybu veslování nahoru. Jak začíná tleskací pohyb, náběžné hrany se setkávají a otáčejí se společně, dokud mezera nezmizí. Zpočátku se předpokládalo, že se křídla dotýkají, ale několik incidentů naznačuje mezeru mezi křídly a naznačuje, že poskytuje aerodynamický přínos.
Ke vzniku výtahu z mechanismu tleskání a mávnutí dochází během několika procesů v celém pohybu. Za prvé, mechanismus se spoléhá na interakci křídlo-křídlo, protože pohyb jednoho křídla nevytváří dostatečný vztlak. Když se křídla pohybují kolem odtokové hrany, vrhá se vzduch do vytvořené mezery a vytváří silný vír na náběžné hraně a druhé se vyvíjí na koncích křídel. Třetí, slabší, vír se vyvíjí na odtokové hraně. Síla rozvíjejících se vírů částečně závisí na počáteční mezeře mezikřídlového oddělení na začátku pohybu. Se sníženou mezerou mezikřídlová mezera indikující větší generaci zdvihu, na úkor větších tažných sil. Bylo zjištěno, že implementace zdvihacího pohybu během házení, pružných křídel a zpožděného blokovacího mechanismu posiluje stabilitu a připevnění víru. Nakonec, aby se kompenzoval celkový nižší vztlak při letu s nízkým Reynoldsovým číslem (s laminárním prouděním ), má drobný hmyz často vyšší frekvenci zdvihů, aby generoval rychlosti křídel, které jsou srovnatelné s větším hmyzem.
Celkově největší očekávané tažné síly se vyskytují během hřbetního pohybu, protože křídla se musí oddělit a otáčet. K útlumu velkých tažných sil dochází několika mechanismy. Bylo zjištěno, že pružná křídla snižují odpor při pohybu křídla až o 50% a dále snižují celkový odpor v celém zdvihu křídla ve srovnání s tuhými křídly. Štětiny na okrajích křídel, jak je vidět na Encarsia formosa , způsobují pórovitost toku, která zvyšuje a snižuje tažné síly, za cenu nižší generace zdvihu. Kromě toho mezikřídlové oddělení před vrhnutím hraje důležitou roli v celkovém účinku odporu. Jak se vzdálenost mezi křídly zvětšuje, celkový odpor klesá.
Mechanismus tleskání a házení využívá také mořský měkkýš Limacina helicina , mořský motýl. Některý hmyz, jako je například listník zeleniny Liriomyza sativae (moucha), využívá částečné tleskání a vrhá se, přičemž mechanismus pouze na vnější části křídla zvyšuje vztlak o přibližně 7% při vznášení.
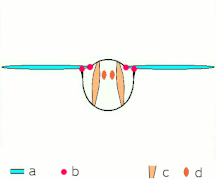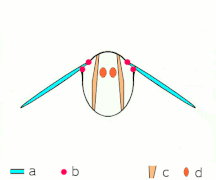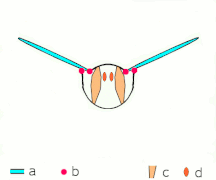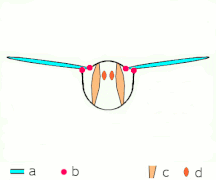
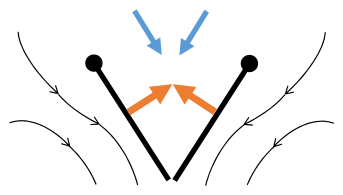
- Clap and fling flight mechanism after Sane 2003

Clap 1: křídla se zavírají nad zády

Clap 2: náběžné hrany se dotýkají, křídlo se otáčí kolem náběžné hrany , tvoří se víry

Clap 3: zadní hrany se zavírají, víry se prolévají, křídla se zavírají a dávají tah



- Černé (zakřivené) šipky: tok; Modré šipky: indukovaná rychlost; Oranžové šipky: čistá síla na křídle
Fling 1: křídla se otáčejí kolem odtokové hrany, aby se rozletěla

Fling 2: náběžná hrana se vzdaluje, proudí vzduch a zvyšuje vztlak
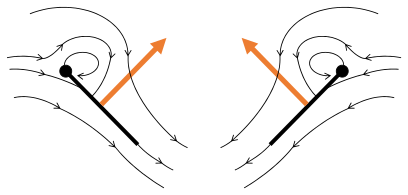
Fling 3: nové vírové formy na náběžné hraně, víry na odtokových hranách se navzájem ruší, možná pomáhají toku rychleji růst ( Weis-Fogh 1973)



Řídící rovnice
Křídlo pohybující se v tekutinách zažívá tekutinovou sílu , která se řídí konvencemi nalezenými v aerodynamice. Silová složka kolmá ke směru toku vzhledem ke křídlu se nazývá zdvih ( L ) a složka síly v opačném směru toku je odpor ( D ). Při uvažovaných Reynoldsových číslech je odpovídající silová jednotka 1/2 (ρU 2 S), kde ρ je hustota tekutiny, S plocha křídla a U rychlost křídla. Bezrozměrné síly se nazývají koeficienty zdvihu ( C L ) a odporu ( C D ), to znamená:

C L a C D jsou konstanty pouze tehdy, je -li tok ustálený. Zvláštní třída předmětů, jako jsou křídla, může dosáhnout ustáleného stavu, když prořízne tekutinu pod malým úhlem náběhu. V tomto případě může být inviscidní tok kolem profilu křídla aproximován potenciálním tokem, který splňuje okrajové podmínky bez penetrace. Kutta-Joukowski věta 2d profilu dále předpokládá, že proud opouští ostrou odtokovou hranou bez problémů, a to určuje celkový krevní oběh kolem profilu. Odpovídající vztlak je dán Bernoulliho zásadou ( Blasiova věta ):

Toky kolem ptáků a hmyzu lze považovat za nestlačitelné : Machovo číslo neboli rychlost vzhledem k rychlosti zvuku ve vzduchu je typicky 1/300 a frekvence křídla je asi 10–103 Hz. Pomocí řídící rovnice jako Navier-Stokesovy rovnice podléhající okrajové podmínce bez prokluzu je rovnice:

Kde u (x, t) je pole toku, p tlak, ρ hustota tekutiny, ν kinematická viskozita, u bd rychlost na hranici a u s rychlost pevné látky. Výběrem délky stupnice, L, a měřítko rychlosti, U, rovnice může být vyjádřena ve formě obsahující bezrozměrný Reynoldsova čísla, R e = ul / vmax. Mezi křídlem hmyzu a křídlem jsou dva zjevné rozdíly : Křídlo hmyzu je mnohem menší a klopí. Na příkladu vážky má akord (c) asi 1 cm (0,39 palce), délku křídla (l) asi 4 cm (1,6 palce) a frekvenci křídla (f) asi 40 Hz. Rychlost hrotu (u) je asi 1 m/s (3,3 ft/s) a odpovídající Reynoldsovo číslo asi 103. Na menším konci má chalcidská vosa délku křídla asi 0,5–0,7 mm ( 0,020–0,028 palce) ) a porazí své křídlo asi na 400 Hz. Jeho Reynoldsovo číslo je asi 25. Rozsah Reynoldsova čísla při letu hmyzu je asi 10 až 10 4 , což leží mezi dvěma limity, které jsou pro teorie vhodné: neviditelné ustálené proudění kolem profilu křídla a Stokesův proud, který zažívá plavecká bakterie. Z tohoto důvodu není tento mezilehlý rozsah dobře srozumitelný. Na druhou stranu je to možná nejvíce všudypřítomný režim mezi věcmi, které vidíme. Padající listy a semena, ryby a ptáci se setkávají s nestálými proudy podobnými tokům kolem hmyzu. Chordwise Reynoldsovo číslo lze popsat takto:

a


Kde je průměrná délka tětivy, rychlost hrotu křídla, amplituda zdvihu, frekvence úderů, poloměr otáčení, plocha křídla a délka křídla včetně špičky křídla.







Kromě Reynoldsova čísla existují ještě další dva relevantní bezrozměrné parametry. Křídlo má tři stupnice rychlosti: klopná rychlost vzhledem k tělu ( u ), dopředná rychlost tělesa ( U 0 ) a stoupající rychlost (Ω c ). Jejich poměry tvoří dvě bezrozměrné proměnné, U 0 / u a Ωc / u , první je často označován jako poměr zálohy, a souvisí také se sníženou frekvencí, fc / U 0 .
Pokud je křídlo hmyzu například tuhé, křídlo Drosophila je přibližně takové, jeho pohyb vzhledem k pevnému tělu lze popsat třemi proměnnými: polohou hrotu v sférických souřadnicích (Θ (t), Φ (t) ) a úhel sklonu ψ (t), kolem osy spojující kořen a špičku. Pro odhad aerodynamických sil na základě analýzy lopatkových prvků je také nutné určit úhel náběhu (α). Typický úhel útoku při 70% rozpětí křídel se pohybuje od 25 ° do 45 ° u vznášejícího se hmyzu (15 ° u kolibříků). Navzdory množství údajů dostupných pro mnoho hmyzu, relativně málo experimentů uvádí časovou změnu α během mrtvice. Patří mezi ně experimenty s uvázaným kobylkou a uvázanou muškou v aerodynamickém tunelu a volný vznášející se let ovocné mušky.
Vzhledem k tomu, že jsou relativně snadno měřitelné, byly trajektorie konců křídel hlášeny častěji. Například výběr pouze letových sekvencí, které vytvářejí dostatečný vztlak na podporu váhy, ukáže, že špička křídla sleduje eliptický tvar. Nekřížené tvary byly hlášeny také u jiného hmyzu. Bez ohledu na jejich přesný tvar, pohyb upínání dolů naznačuje, že hmyz může kromě zvedání používat také aerodynamický odpor, aby podpořil svou váhu.
Vznášející se
| Letové parametry | Rychlost (m/s) | Beats/s |
|---|---|---|
| Vážka Aeshnid | 7.0 | 38 |
| Sršeň | 5.7 | 100 |
| Kolibří hawkmoth | 5,0 | 85 |
| Ovád | 3.9 | 96 |
| Vznášedlo | 3.5 | 120 |
| Čmelák | 2.9 | 130 |
| Včelí med | 2.5 | 250 |
| Moucha | 2.0 | 190 |
| Motýlí | 1.5 | 16 |
| Scorpionfly | 0,49 | 28 |
| Velký bílý motýl | 2.5 | 12 |
| Thrips (tleskání a mávnutí) | 0,3 | 254 |
.jpg)
Mnoho hmyzu se může vznášet nebo zůstat na jednom místě ve vzduchu, a to tak, že rychle porazí křídla. K tomu je nutná boční stabilizace a výroba výtahu. Zvedací síla je produkována hlavně dolů. Jak křídla tlačí dolů na okolní vzduch, výsledná reakční síla vzduchu na křídlech tlačí hmyz nahoru. Křídla většiny hmyzu jsou vyvinuta tak, aby během zdvihu nahoru byla síla na křídlo malá. Vzhledem k tomu, že nárazový a zpětný zdvih nutí hmyz nahoru a dolů, hmyz osciluje a končí ve stejné poloze.
Vzdálenost hmyzu mezi údery křídel závisí na tom, jak rychle jeho křídla bijí: čím pomaleji klapá, tím delší je interval, ve kterém padá, a čím dále klesá mezi jednotlivými křídly. Lze vypočítat frekvenci úderů křídel nezbytnou k tomu, aby si hmyz udržel danou stabilitu ve své amplitudě. Pro zjednodušení výpočtů je třeba předpokládat, že zvedací síla je na konečné konstantní hodnotě, když se křídla pohybují dolů, a že je nulová, když se křídla pohybují nahoru. Během časového intervalu Δ t vzestupného úderu křídla hmyz klesne pod vlivem gravitace na vzdálenost h .

Zdvih nahoru pak vrátí hmyz do původní polohy. Typicky může být požadováno, aby se svislá poloha hmyzu nezměnila o více než 0,1 mm (tj. H = 0,1 mm). Maximální povolená doba pro volný pád je pak

Protože pohyby nahoru a dolů křídel trvají přibližně stejně dlouho, je perioda T pro úplné křídlo nahoru a dolů dvakrát Δ r , tj.

Frekvence úderů, f, což znamená počet křídel za sekundu, je reprezentována rovnicí:

V použitých příkladech je použitá frekvence 110 tepů/s, což je typická frekvence u hmyzu. Motýli mají mnohem pomalejší frekvenci přibližně 10 úderů/s, což znamená, že se nemohou vznášet. Jiný hmyz může být schopen produkovat frekvenci 1000 úderů/s. Aby se hmyz vrátil do původní svislé polohy, musí se průměrná síla vzhůru během zdvihu dolů F av rovnat dvojnásobku hmotnosti hmyzu. Všimněte si toho, protože vzhledem k tomu, že síla vzhůru na tělo hmyzu působí pouze polovinu času, průměrná síla vzhůru na hmyz je jednoduše jeho hmotnost.
Příkon
Nyní lze vypočítat sílu potřebnou k udržení vznášení, přičemž znovu uvažujeme o hmyzu s hmotností m 0,1 g, průměrná síla, F av , aplikovaná oběma křídly během zdvihu dolů, je dvojnásobek hmotnosti. Vzhledem k tomu, že tlak působící na křídla je rovnoměrně rozložen po celé ploše křídla, znamená to, že lze předpokládat, že síla generovaná každým křídlem působí jediným bodem v polovině křídel. Během zdvihu směrem dolů prochází střed křídel svislou vzdáleností d . Celková práce, kterou hmyz vykonal během každého tahu dolů, je součinem síly a vzdálenosti; to znamená,

Pokud se křídla otáčí úderem pod úhlem 70 °, pak v případě uvedeném pro hmyz s křídly dlouhými 1 cm je d 0,57 cm. Práce odvedená během každého zdvihu dvěma křídly je tedy:

Energie se používá ke zvýšení hmyzu proti gravitaci. Energie E potřebná ke zvýšení hmotnosti hmyzu o 0,1 mm při každém sestupu je:

To je zanedbatelný zlomek celkové vynaložené energie, což zjevně znamená, že většina energie je vynaložena v jiných procesech. Podrobnější analýza problému ukazuje, že práce odvedená křídly je přeměněna primárně na kinetickou energii vzduchu, která je zrychlena pohybem křídel směrem dolů. Výkon je množství práce provedené za 1 s; u hmyzu použitého jako příklad provede 110 tahů dolů za sekundu. Jeho výstupní výkon P je tedy zdvihy za sekundu, a to znamená, že jeho výstupní výkon P je:

Výkon
Při výpočtu síly použité při vznášení zanedbaly použité příklady kinetickou energii pohybujících se křídel. Křídla hmyzu, jak jsou lehká, mají konečnou hmotnost; proto, když se pohybují, mají kinetickou energii. Protože jsou křídla v rotačním pohybu, maximální kinetická energie při každém zdvihu křídla je:

Zde I je moment setrvačnosti křídla a ω max je maximální úhlová rychlost během zdvihu křídla. Abychom získali moment setrvačnosti pro křídlo, budeme předpokládat, že křídlo lze aproximovat tenkou tyčí otočnou na jednom konci. Moment setrvačnosti křídla je pak:

Kde l je délka křídla (1 cm) a m je hmotnost dvou křídel, která může být obvykle 10-3 g. Maximální úhlovou rychlost ω max lze vypočítat z maximální lineární rychlosti ν max ve středu křídla:

Během každého zdvihu se střed křídel pohybuje průměrnou lineární rychlostí ν av danou vzdáleností d uraženou středem křídla dělenou dobou Δt zdvihu křídla. Z našeho předchozího příkladu d = 0,57 cm a Δt = 4,5 × 10 −3 s. Proto:

Rychlost křídel je nulová jak na začátku, tak na konci zdvihu křídla, což znamená, že maximální lineární rychlost je vyšší než průměrná rychlost. Pokud předpokládáme, že rychlost osciluje ( sinusově ) po dráze křídla, je maximální rychlost dvakrát vyšší než průměrná rychlost. Maximální úhlová rychlost je tedy:

A kinetická energie je tedy:

Protože v každém cyklu pohybu křídla existují dva zdvihy křídla (zdvih nahoru a dolů), je kinetická energie 2 × 43 = 86 erg . To je asi tolik energie, kolik se spotřebuje při samotném vznášení.
Pružnost

Hmyz získává kinetickou energii zajišťovanou svaly, když křídla zrychlují . Když křídla začnou ke konci tahu zpomalovat, musí se tato energie rozptýlit. Při sestupu je kinetická energie rozptýlena samotnými svaly a přeměněna na teplo (toto teplo se někdy používá k udržení tělesné teploty). Některý hmyz je schopen využít kinetickou energii při pohybu křídel vzhůru, aby jim pomohl při letu. Křídlové klouby tohoto hmyzu obsahují polštářek z elastického gumového proteinu zvaného pryskyřice . Během zdvihu křídla se pryskyřice natáhne. Kinetická energie křídla je přeměněna na potenciální energii v nataženém pryskyřici, která energii ukládá podobně jako pružina. Když se křídlo pohybuje dolů, tato energie se uvolňuje a pomáhá při stahování dolů.
Pomocí několika zjednodušujících předpokladů můžeme vypočítat množství energie uložené v natažené pryskyřici. Přestože je pryskyřice ohnuta do složitého tvaru, uvedený příklad ukazuje výpočet jako přímou tyč o ploše A a délce. Dále budeme předpokládat, že v celém úseku se pryskyřice řídí Hookeovým zákonem . To není úplně pravda, protože pryskyřice je natažena o značné množství, a proto se v procesu roztahování mění jak plocha, tak Youngův modul . Potenciální energie U uložená v natažené pryskyřici je:

Zde E je Youngův modul pro resilin, který byl změřen na 1,8 x 10 7 dyn / cm 2 . Typicky v hmyzu velikosti včela, objem resilin může být ekvivalentní k válci 2 x 10 -2 cm a 4 x 10 -4 cm 2 v oblasti. V uvedeném příkladu je délka pryskyřičného prutu prodloužena o 50%. To znamená, že Δℓ je 10 -2 cm. V tomto případě je tedy potenciální energie uložená v pryskyřici každého křídla:

Uložená energie ve dvou křídlech pro hmyz velikosti včel je 36 erg, což je srovnatelné s kinetickou energií při zdvihu křídel. Experimenty ukazují, že až 80% kinetické energie křídla může být uloženo v pryskyřici.
Křídlová spojka

Některé čtyřkřídlé hmyzí řády, jako například Lepidoptera, vyvinuly v imagu morfologické spojovací mechanismy křídel, které činí tyto taxony funkčně dvoukřídlými. Všechny kromě nejzákladnějších forem vykazují tuto křídlovou spojku.
Mechanismy jsou tří různých typů - jugalské, frenulo -retinakulární a amplexiformní:
- Primitivnější skupiny mají zvětšenou lalokovitou oblast poblíž bazálního zadního okraje, tj. Na bázi předního křídla, a jugum , které se za letu složí pod zadní křídlo .
- Jiné skupiny mají na zadním křídle frenulum, které se na předním křídle zavěšuje pod retinaculum.
- Téměř ve všech motýlech a v Bombycoidea (kromě Sphingidae ) neexistuje žádné uspořádání frenulum a retinaculum pro spojení křídel. Místo toho je rozšířená humerální oblast zadního křídla široce překryta předním křídlem. Navzdory absenci specifického mechanického spojení se křídla překrývají a pracují ve fázi. Silový zdvih předního křídla souběžně tlačí dolů zadní křídlo. Tento typ spojky je variací frenátového typu, ale kde jsou frenulum a retinaculum zcela ztraceny.
Biochemie
Biochemie letu hmyzu byla předmětem značné studie. Zatímco mnoho hmyzu používá jako zdroj energie k letu sacharidy a lipidy , mnoho brouků a much používá jako zdroj energie aminokyselinu prolin . Některé druhy také používají kombinaci zdrojů a můry, jako například Manduca sexta, používají uhlovodany k předletové rozcvičce.
Evoluce a adaptace

Někdy v karbonském období , asi před 350 až 400 miliony let, kdy existovaly pouze dvě hlavní pevniny, začal létat hmyz. Mezi nejstarší zkřídlé hmyzí zkameněliny patří Delitzschala , paleodiktoptéra ze spodního karbonu; Rhyniognatha je starší, z raného devonu , ale není jisté, zda měla křídla, nebo byl skutečně hmyzem.
Jak a proč se vyvinula křídla hmyzu, není dobře pochopeno, a to především z důvodu nedostatku vhodných zkamenělin z období jejich vývoje v spodním karbonu. Historicky existovaly tři hlavní teorie o původu letu hmyzu. První byl, že se modifikace pohyblivých břišních žáber , jak bylo zjištěno na vodní naiads z jepice . Fylogenomická analýza naznačuje, že Polyneoptera , skupina okřídleného hmyzu, která zahrnuje kobylky, se vyvinula z pozemského předka, takže vývoj křídel ze žábry je nepravděpodobný. Další studie skokového chování larev jepice zjistila, že tracheální žábry nehrají žádnou roli při vedení hmyzího původu, což poskytuje další důkazy proti této evoluční hypotéze. Zbývají tedy dvě hlavní historické teorie: že křídla se vyvinula z paranotálních laloků, prodloužení hrudní tergy ; nebo že vznikly z modifikací segmentů nohou, které již obsahovaly svaly.
Hypotéza epikoxální (břišní žábry)
.JPG)
Četné entomologové včetně Landois v roce 1871, Lubbock v roce 1873, Graber v roce 1877, a Osborna v roce 1905 naznačují, že možný původ pro hmyzí křídla mohla být movité břišní žábry nalézt v mnoha vodní hmyz, například na naiads z jepice. Podle této teorie se tyto tracheální žábry, které začaly svou cestou jako východy z dýchacího ústrojí a postupem času byly upraveny na lokomotivní účely, nakonec vyvinuly v křídla. Tracheální žábry jsou vybaveny malými křidélky, která neustále vibrují a mají vlastní drobné rovné svaly.
Paranotální (tergální) hypotéza
Hypotéza paranotálního laloku nebo tergalu (hřbetní tělesná stěna), navržená Fritzem Müllerem v roce 1875 a přepracovaná G. Cramptonem v roce 1916, Jarmilou Kulakovou-Peckovou v roce 1978 a Alexandrem P. Rasnitsynem v roce 1981, mimo jiné naznačuje, že křídla hmyzu se vyvinula z paranotální laloky, předběžná adaptace nalezená ve zkamenělinách hmyzu, která by napomohla stabilizaci při skákání nebo padání. Ve prospěch této hypotézy je tendence většiny hmyzu, když se při lezení na větvích lekne, uniknout pádem na zem. Takové laloky by sloužily jako padáky a umožnily hmyzu přistát měkčeji. Teorie naznačuje, že tyto laloky se postupně zvětšovaly a v pozdější fázi se vytvořil kloub s hrudníkem. Ještě později se objevily svaly pro pohyb těchto hrubých křídel. Tento model znamená postupné zvyšování účinnosti křídel, počínaje padákem , poté klouzáním a nakonec aktivním letem . Přesto nedostatek zásadních fosilních důkazů o vývoji křídlových kloubů a svalů představuje pro teorii zásadní problém, stejně jako zdánlivě spontánní vývoj artikulace a žilnatosti, a to odborníci v oboru do značné míry odmítli.
Endite-exite (pleurální) hypotéza

V roce 1990 JWH Trueman navrhl, aby křídlo bylo adaptováno z enditů a exitů , příloh o příslušných vnitřních a vnějších aspektech primitivní končetiny členovců , nazývaných také pleurální hypotéza. Toto bylo založeno na studii Goldschmidta v roce 1945 na Drosophila melanogaster , ve které variace nazvaná "pod" (pro podomeres, segmenty končetin) vykazovala mutaci, která transformovala normální křídla. Výsledek byl interpretován jako trojité kloubové uspořádání nohou s některými dalšími přídavky, ale bez tarzu, kde by normálně byl pobřežní povrch křídla. Tato mutace byla reinterpretována jako silný důkaz pro dorzální exite a endite fusion, spíše než pro nohu, přičemž přívěsky zapadají do této hypotézy mnohem lépe. Inervace, artikulace a svalstvo potřebné pro vývoj křídel jsou již přítomny v končetinových segmentech.
Jiné hypotézy
Mezi další hypotézy patří návrh Vincenta Wiggleswortha z roku 1973, že křídla se vyvinula z hrudních výčnělků používaných jako radiátory .
Adrian Thomas a Åke Norberg v roce 2003 navrhli, že křídla se mohla původně vyvinout pro plavbu na povrchu vody, jak je vidět na některých kamenných muškách .
Stephen P. Yanoviak a jeho kolegové v roce 2009 navrhli, aby křídlo pocházelo z přímého sestupového leteckého sestupu - fenomén předletu , který se vyskytuje u nějaké apterygoty , bezkřídlého sesterského taxonu pro okřídlený hmyz.
Duální původ
Biologové včetně Averofa, Niwy, Eliase-Neta a jejich kolegů začali kromě paleontologických důkazů zkoumat původ křídla hmyzu pomocí evo-devo . To naznačuje, že křídla jsou sériově homologní s tergálními i pleurálními strukturami, což potenciálně vyřeší staletou debatu. Jakub Prokop a jeho kolegové našli v roce 2017 paleontologické důkazy z paleozoických nymfových křídel, že křídla měla skutečně dvojí původ.
Viz také
Reference
Další čtení
- Dickinson, MH; Lehmann, FO; Sane, SP (1999). „Rotace křídla a aerodynamický základ letu hmyzu“. Věda . 284 (5422): 1954–60. doi : 10,1126/věda.284.5422.1954 . PMID 10373107 .
- Ellington, Charles P .; Van Den Berg, Coen; Willmott, Alexander P .; Thomas, Adrian LR (1996). „Špičkové víry při letu hmyzu“. Příroda . 384 (6610): 626–630. Bibcode : 1996Natur.384..626E . doi : 10,1038/384626a0 . S2CID 4358428 .
- Ellington, CP (1984). „Aerodynamika vznášení hmyzu. I. Kvazi-ustálená analýza“ . Filozofické transakce Královské společnosti B: Biologické vědy . 305 (1122): 1–15. Bibcode : 1984RSPTB.305 .... 1E . doi : 10,1098/rstb.1984.0049 .
- Ellington, CP (1999). „Nová aerodynamika letu hmyzu: Aplikace pro mikro-vzduchová vozidla“ . The Journal of Experimental Biology . 202 (Pt 23): 3439–3448. doi : 10,1242/jeb.202.23.3439 . PMID 10562527 .
- Lewin, GC; Haj-Hariri, H. (2003). „Modelování tahové generace dvourozměrného zvedaného profilu křídla ve viskózním proudění“. Journal of Fluid Mechanics . 492 : 339–362. Bibcode : 2003JFM ... 492..339L . doi : 10,1017/S0022112003005743 . S2CID 122077834 .
- Pringle, JWS (1975). Let hmyzem . Oxfordští čtenáři biologie. 52 .
- Sane, SP (2003). „Aerodynamika letu hmyzu“ . The Journal of Experimental Biology . 206 (Pt 23): 4191–208. doi : 10,1242/jeb.00663 . PMID 14581590 .
- Savage, SB; Newman, BG; Wong, DTM (1979). „Role vírů a nestálých efektů během vznášejícího se letu dračích much“ . The Journal of Experimental Biology . 83 (1): 59–77. doi : 10,1242/jeb.83.1.59 .
- Smyth, T., Jr. (1985). „Svalové systémy“. In Blum, MS Blum (ed.). Základy fyziologie hmyzu . John Wiley and Sons. s. 227–286.
- Sotavalta, O. (1953). „Záznamy vysokého zdvihu křídla a hrudní vibrační frekvence u některých midges“ . Biol. Býk. Woods Hole . 104 (3): 439–444. doi : 10,2307/1538496 . JSTOR 1538496 .
- Van Den Berg, C .; Ellington, CP (1997). „Vírová probuzení„ vznášejícího se “modelu jestřába“ . Filozofické transakce Královské společnosti B: Biologické vědy . 352 (1351): 317–328. Bibcode : 1997RSPTB.352..317V . doi : 10,1098/rstb.1997.0023 . PMC 1691928 .
- Walker, JA (září 2002). „Rotační zdvih: rozdíl nebo něco podobného?“ . The Journal of Experimental Biology . 205 (Pt 24): 3783–3792. doi : 10,1242/jeb.205.24.3783 . PMID 12432002 .
- Zbikowski, R. (2002). „O aerodynamickém modelování hmyzu podobného třepetajícího se křídla ve vznášedle pro mikro vzdušná vozidla“. Filozofické transakce Královské společnosti A: Matematické, fyzikální a inženýrské vědy . 360 (1791): 273–290. Bibcode : 2002RSPTA.360..273Z . doi : 10.1098/rsta.2001.0930 . PMID 16210181 . S2CID 2430367 .
{kind=link}