
Pilus - Pilus

Pilus ( latina pro ‚vlasy‘; množné číslo: pili ) je vlasy jako přívěsek nalezen na povrchu mnoha bakterií a archeí . Termíny pilus a fimbria (latinsky 'fringe'; množné číslo: fimbriae ) lze použít zaměnitelně, ačkoli někteří badatelé si vyhrazují termín pilus pro přívěsek požadovaný pro bakteriální konjugaci . Všechny pili v druhém smyslu jsou primárně složeny z pilinových proteinů, které jsou oligomerní .
Na bakteriálním a archaealním povrchu mohou existovat desítky těchto struktur. Některé bakterie, viry nebo bakteriofágy se připojují k receptorům na pili na začátku jejich reprodukčního cyklu.
Pili jsou antigenní . Jsou také křehké a neustále nahrazovány, někdy piliemi různého složení, což vede ke změně antigenicity. Specifické reakce hostitele na starou strukturu pili nejsou na novou strukturu účinné. Rekombinační geny pili kódují variabilní (V) a konstantní (C) oblasti pili (podobné diverzitě imunoglobulinů ). Jako primární antigenní determinanty, faktory virulence a faktory beztrestnosti na buněčném povrchu řady druhů gramnegativních a některých grampozitivních bakterií, včetně Enterobacteriaceae , Pseudomonadaceae a Neisseriaceae , existuje velký zájem o studium pili jako organel adheze a jako složky vakcíny. První podrobnou studii pili provedli Brinton a spolupracovníci, kteří prokázali existenci dvou odlišných fází v rámci jednoho bakteriálního kmene: pileated (p+) a non-pileated)
Typy podle funkce
Několik jmen je dáno různým typům pili podle jejich funkce. Klasifikace se vždy nepřekrývá se strukturálními nebo evolučními typy, protože dochází ke konvergentní evoluci .
Konjugativní pili
Konjugativní pili umožňují přenos DNA mezi bakteriemi v procesu bakteriální konjugace . Někdy se jim říká „sex pili“, analogicky k sexuální reprodukci , protože umožňují výměnu genů prostřednictvím tvorby „párových párů“. Asi nejlépe prozkoumaný je F-pilus Escherichia coli , kódovaný F sexuálním faktorem .
Sex pilus má obvykle průměr 6 až 7 nm . Během konjugace pilus vycházející z dárcovské bakterie polapí přijímající bakterii, přitáhne ji k sobě a nakonec spustí tvorbu pářícího můstku , který naváže přímý kontakt a tvorbu kontrolovaného póru, který umožňuje přenos DNA z dárce do příjemce. Přenesená DNA se obvykle skládá z genů potřebných k výrobě a přenosu pili (často kódovaných na plazmidu ), a tak je to také druh sobecké DNA ; často se však společně přenášejí i jiné kousky DNA, což může vést k šíření genetických znaků v bakteriální populaci, jako je rezistence na antibiotika . Ne všechny bakterie mohou vytvářet konjugační pili, ale konjugace může nastat mezi bakteriemi různých druhů.
Fimbriae
Fimbria je termín používaný pro krátký pilus, který se používá k přichycení bakterie k povrchu, někdy se mu také říká „připevňovací pilus“. Fimbrie jsou nezbytné pro tvorbu biofilmu, protože připojují bakterie k hostitelským povrchům pro kolonizaci během infekce. Fimbrie jsou buď umístěny na pólech buňky, nebo jsou rovnoměrně rozloženy po celém jejím povrchu. Mutantní bakterie, které postrádají fimbrie, se nemohou přichytit ke svým obvyklým cílovým povrchům, a nemohou tedy způsobit nemoci .
Tento termín byl také použit v laxním smyslu pro označení všech pili, těmi, kteří používali „pilus“ konkrétně pro sex pili.
Typy sestavením systému nebo struktury
Převod
Rodina Tra (převod) zahrnuje všechny známé sex pili (od roku 2010). Jsou příbuzné sekrečnímu systému typu IV (T4SS). Mohou být zařazeny do typu F (po F-pilusu) a typu P. Pilus, stejně jako jejich sekreční protějšky, vstřikuje materiál, v tomto případě DNA, do jiné buňky.
Pili typu IV

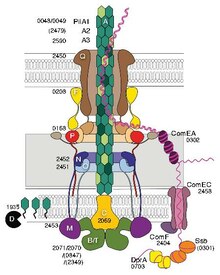
Některé pilíře, nazývané pili typu IV (T4P), vytvářejí pohyblivé síly. Vnější konce pilíře přilnou k pevnému substrátu, buď k povrchu, ke kterému je bakterie připojena, nebo k jiným bakteriím. Když se pili stáhnou, vytáhnou bakterii dopředu jako hák. Pohyb produkovaný pili typu IV je typicky trhaný, takže se nazývá záškubová motilita , na rozdíl od jiných forem bakteriální motility, jako je ta, kterou produkují bičíky . Některé bakterie, například Myxococcus xanthus , však vykazují klouzavou pohyblivost . Bakteriální pili typu IV mají podobnou strukturu jako komponent flagellinu z archaella (archaeal flagella), a oba jsou spojeny s sekreční systém typu II (T2SS); jsou sjednoceny skupinou filamentových systémů typu IV . Kromě archaelly mnoho archaea produkuje adhezivní pili typu 4, které umožňují archaeálním buňkám přilnout k různým substrátům. N-koncové alfa-šroubovicové části archaealních pilířů typu 4 a archaellinů jsou homologní s odpovídajícími oblastmi bakteriálního T4P; zdá se však, že domény bohaté na C-koncový beta řetězec nesouvisejí v bakteriálních a archaealních pilířích.
Genetická transformace je proces, při kterém hostitelská bakteriální buňka odebere DNA ze sousední buňky a integruje tuto DNA do svého genomu homologní rekombinací . U Neisseria meningitidis (také nazývaného meningococcus) vyžaduje transformace DNA přítomnost krátkých sekvencí vychytávání DNA (DUS), což je 9-10 monomerů sídlících v kódujících oblastech dárcovské DNA. Zvláštní uznání Duss je zprostředkována typu IV Pilin . Pili Menningococcal typu IV vážou DNA prostřednictvím menšího pilinu ComP prostřednictvím elektropozitivního pruhu, o kterém se předpovídá, že bude vystaven na povrchu vlákna. ComP zobrazuje vynikající preference vazby pro selektivní DUS. Distribuce DUS v genomu N. meningitides zvýhodňuje určité geny, což naznačuje, že existuje zaujatost pro geny podílející se na údržbě a opravách genomu.
Tato rodina byla původně identifikována jako „fimbrie typu IV“ podle jejich vzhledu pod mikroskopem. Tato klasifikace přežila, protože shodou okolností odpovídá kladu.
Virulence
Pili jsou zodpovědné za virulenci v patogenních kmenech mnoha bakterií, včetně E. coli , Vibrio cholerae a mnoha kmenů Streptococcus . Důvodem je, že přítomnost pili výrazně zvyšuje schopnost bakterií vázat se na tělesné tkáně, což pak zvyšuje rychlost replikace a schopnost interakce s hostitelským organismem. Pokud má určitý druh bakterie více kmenů, ale pouze některé jsou patogenní, je pravděpodobné, že patogenní kmeny budou mít pili, zatímco nepatogenní kmeny nebudou.
Rozvoj připojovacího pilíře pak může vést k rozvoji dalších znaků virulence. Nepatogenní kmeny V. cholerae nejprve vyvinuly pili, což jim umožnilo vázat se na lidské tkáně a vytvářet mikrokolonie . Tyto pili pak sloužily jako vazebná místa pro lysogenní bakteriofág, který nese toxin způsobující onemocnění . Gen pro tento toxin, jakmile je začleněn do genomu bakterie, je exprimován, když je exprimován gen kódující pilus (odtud název „pilin zprostředkovaný toxinem“).
Viz také
Reference
externí odkazy
- Sex+Pilus v Americké národní lékařské knihovně Předměty (MeSH)
- Bakteriální+Pilus v USA Národní knihovna lékařských lékařských oborových nadpisů (MeSH)