
Neproteinogenní aminokyseliny - Non-proteinogenic amino acids

V biochemii , non-kódované nebo neproteinogenních aminokyselin se liší od 22 proteinogenních aminokyselin (21 u eukaryot), které jsou přirozeně kódovaných v genomu organismů pro montáž proteinů. V bílkovinách se však přirozeně vyskytuje přes 140 neproteinogenních aminokyselin a tisíce dalších se mohou vyskytovat v přírodě nebo být syntetizovány v laboratoři. Důležité je mnoho neproteinogenních aminokyselin:
- meziprodukty v biosyntéze,
- při posttranslační tvorbě proteinů,
- ve fyziologické roli (např. složky stěn bakteriálních buněk , neurotransmitery a toxiny ),
- přírodní nebo člověkem vyrobené farmakologické sloučeniny,
- přítomné v meteoritech nebo používané v prebiotických experimentech (např. Miller – Ureyův experiment ).
Definice negací

Technicky, jakákoli organická sloučenina s aminem (-NH 2 ) a karboxylové kyseliny (-COOH) funkční skupinu je aminokyselina. Proteinogenní aminokyseliny jsou malou podskupinou této skupiny, která má centrální atom uhlíku (α- nebo 2-) nesoucí aminoskupinu, karboxylovou skupinu, postranní řetězec a α-vodíkovou levo konformaci , s výjimkou glycinu , který je achirální a prolin , jejichž aminová skupina je sekundární amin, a proto je z tradičních důvodů často označována jako imino kyselina , i když ne imino.
Genetický kód kóduje 20 standardních aminokyselin pro začlenění do proteinů během translace . Existují však dvě extra proteinogenní aminokyseliny: selenocystein a pyrrolysin . Tyto nestandardní aminokyseliny nemají vyhrazený kodon, ale jsou přidány místo stop kodonu, pokud je přítomna specifická sekvence, UGA kodon a SECIS prvek pro selenocystein, UAG PYLIS downstream sekvence pro pyrrolysin. Všechny ostatní aminokyseliny se označují jako „neproteinogenní“.

Selenocystein. Tato aminokyselina obsahuje na svém β-uhlíku selenolovou skupinu

Pyrrolysin. Tato aminokyselina vzniká spojením karboxylovaného pyrrolinového kruhu s e-aminoskupinou lysinu

Existují různé skupiny aminokyselin:
- 20 standardních aminokyselin
- 22 proteinogenních aminokyselin
- více než 80 aminokyselin vytvořených abioticky ve vysokých koncentracích
- asi 900 je produkováno přírodními cestami
- do proteinu bylo umístěno více než 118 upravených aminokyselin
Tyto skupiny se překrývají, ale nejsou totožné. Všech 22 proteinogenních aminokyselin je biosyntetizováno organismy a některé, ale ne všechny, jsou také abiotické (nalezeno v prebiotických experimentech a meteoritech). Některé přírodní aminokyseliny, jako je norleucin , jsou translinicky chybně začleněny do proteinů kvůli nevěře v proces syntézy proteinů. Mnoho aminokyselin, jako je ornithin , jsou metabolické meziprodukty vyráběné biosynteticky, ale nejsou translativně transponovány do proteinů. Posttranslační modifikace aminokyselinových zbytků v proteinech vede k tvorbě mnoha proteinových, ale neproteinogenních aminokyselin. Jiné aminokyseliny se nacházejí pouze v abiotických směsích (např. Α-methylnorvalin). Více než 30 nepřirozených aminokyselin bylo translačně vloženo do proteinu v inženýrských systémech, ale nejsou biosyntetické.
Nomenklatura
Kromě systému číslování IUPAC k diferenciaci různých uhlíků v organické molekule, postupným přiřazením čísla každému uhlíku, včetně těch, které tvoří karboxylovou skupinu, mohou být uhlíky podél postranního řetězce aminokyselin také označeny řeckými písmeny , kde a-uhlík je centrální chirální uhlík obsahující karboxylovou skupinu, postranní řetězec a v a-aminokyselinách aminoskupinu-uhlík v karboxylových skupinách se nepočítá. (V důsledku toho názvy IUPAC mnoha neproteinogenních α-aminokyselin začínají 2-amino- a končí kyselinou -ic .)
Přírodní, ale ne L-α-aminokyseliny
Většina přírodních aminokyselin jsou α-aminokyseliny v L konformaci, ale existují určité výjimky.
Non-alfa

Některé non-α aminokyseliny existují v organismech. V těchto strukturách se aminová skupina dále vytlačuje z konce karboxylové kyseliny v molekule aminokyseliny. P -aminokyselina má tedy aminoskupinu navázanou na druhý uhlík pryč a yaminokyselina ji má na třetím. Příklady zahrnují p-alanin , GABA a 5- aminolevulinovou kyselinu .

β-alanin: aminokyselina produkovaná aspartát 1-dekarboxylázou a prekurzor koenzymu A a peptidů karnosinu a anserinu .

Kyselina γ-aminomáselná (GABA): neurotransmiter u zvířat.

Kyselina 4-aminobenzoová (PABA): meziprodukt v biosyntéze folátu



Důvod, proč se v bílkovinách používají α-aminokyseliny, je spojen s jejich četností v meteoritech a prebiotických experimentech. Počáteční spekulace o škodlivých vlastnostech beta-aminokyselin z hlediska sekundární struktury se ukázaly jako nesprávné.
D-aminokyseliny
Některé aminokyseliny obsahují opačnou absolutní chiralitu, chemikálie, které nejsou dostupné z normálního ribozomálního translačního/transkripčního zařízení. Většinu stěn bakteriálních buněk tvoří peptidoglykan , polymer složený z aminosacharidů zesítěných krátkými oligopeptidy přemostěnými mezi sebou. Oligopeptid není syntetizován ribozomálně a obsahuje několik zvláštností, včetně D-aminokyselin , obecně D-alaninu a D-glutamátu. Další zvláštnost je to, že první z nich je racemizovat prostřednictvím PLP vážící enzymů (kódovaného ALR nebo homologu DADX ), zatímco druhá je racemizovat pomocí nezávislého enzymový kofaktor ( Muri ). Existují některé varianty, v Thermotoga spp. Je přítomen D-lysin a v některých bakteriích rezistentních na vankomycin je přítomen D-serin ( gen vanT ).
U zvířat jsou některé D-aminokyseliny neurotransmitery.
Bez vodíku na α-uhlíku
Všechny proteinogenní aminokyseliny mají na α-uhlíku alespoň jeden vodík. Glycin má dva vodíky a všechny ostatní mají jeden vodík a jeden postranní řetězec. Nahrazení zbývajícího vodíku větším substituentem, jako je methylová skupina, narušuje proteinovou páteř.
U některých hub se kyselina a-aminoisomáselná vyrábí jako prekurzor peptidů, z nichž některé vykazují antibiotické vlastnosti. Tato sloučenina je podobná alaninu, ale má na a-uhlíku místo vodíku další methylovou skupinu. Je tedy achirální. Další sloučeninou podobnou alaninu bez a-vodíku je dehydroalanin , který má methylenový postranní řetězec. Je to jedna z několika přirozeně se vyskytujících dehydroaminokyselin .

alanin

kyselina aminoisomáselná

dehydroalanin



Dvojitá aminokyselinová stereocentra
Podskupina L-α-aminokyselin je nejednoznačná, pokud jde o to, který ze dvou konců je α-uhlík. V proteinech může cysteinový zbytek tvořit disulfidovou vazbu s jiným cysteinovým zbytkem, čímž se protein zesíťuje. Dva zesítěné cysteiny tvoří molekulu cystinu . Cystein a methionin jsou obecně vyráběny přímou sulfurylací, ale u některých druhů mohou být produkovány transsulfurací , kde je aktivovaný homoserin nebo serin fúzován s cysteinem nebo homocysteinem tvořícím cystathionin . Podobnou sloučeninou je lanthionin , který lze vidět jako dvě molekuly alaninu spojené thioetherovou vazbou a nachází se v různých organismech. Podobně je kyselina djenkolová , rostlinný toxin z fazolí jengkol , složena ze dvou cysteinů spojených methylenovou skupinou. Kyselina diaminopimelová se používá jako můstek v peptidoglykanu a používá se jako prekurzor lysinu (prostřednictvím jeho dekarboxylace).

cystin
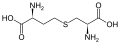
cystathionin

lanthionin
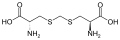
Kyselina djenkolová
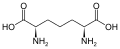
Kyselina diaminopimelová

Prebiotické aminokyseliny a alternativní biochemie
V meteoritech a v prebiotických experimentech (např. Miller -Ureyův experiment ) se nachází mnohem více aminokyselin než dvacet standardních aminokyselin, z nichž několik je ve vyšších koncentracích než standardní: předpokládalo se, že pokud by vznikl život na bázi aminokyselin souběžně jinde ve vesmíru by nebylo společných více než 75% aminokyselin. Nejpozoruhodnější anomálií je nedostatek kyseliny aminomáselné.
| Podíl aminokyselin vzhledem ke glycinu (%) | ||
|---|---|---|
| Molekula | Elektrický výboj | Murchinsonův meteorit |
| glycin | 100 | 100 |
| alanin | 180 | 36 |
| kyselina α-amino-n-máselná | 61 | 19 |
| norvaline | 14 | 14 |
| valin | 4.4 | |
| norleucin | 1.4 | |
| leucin | 2.6 | |
| isoleucin | 1.1 | |
| aloisoleucin | 1.2 | |
| t-leucin | <0,005 | |
| kyselina α-amino-n-heptanová | 0,3 | |
| prolin | 0,3 | 22 |
| pipekolová kyselina | 0,01 | 11 |
| kyselina α, β-diaminopropionová | 1.5 | |
| kyselina α, γ-diaminomáselná | 7.6 | |
| ornitin | <0,01 | |
| lysin | <0,01 | |
| kyselina asparagová | 7.7 | 13 |
| kyselina glutamová | 1.7 | 20 |
| serin | 1.1 | |
| threonin | 0,2 | |
| alothreonin | 0,2 | |
| methionin | 0,1 | |
| homocystein | 0,5 | |
| homoserine | 0,5 | |
| β-alanin | 4.3 | 10 |
| kyselina p-amino-n-máselná | 0,1 | 5 |
| kyselina β-aminoisomáselná | 0,5 | 7 |
| kyselina y-aminomáselná | 0,5 | 7 |
| kyselina α-aminoisomáselná | 7 | 33 |
| isovalin | 1 | 11 |
| sarkosin | 12.5 | 7 |
| N-ethylglycin | 6.8 | 6 |
| N-propylglycin | 0,5 | |
| N-isopropylglycin | 0,5 | |
| N-methylalanin | 3.4 | 3 |
| N-ethyl alanin | <0,05 | |
| N-methyl-beta-alanin | 1,0 | |
| N-ethyl β-alanin | <0,05 | |
| isoserin | 1.2 | |
| kyselina a-hydroxy-y-aminomáselná | 17 |
Přímý boční řetěz
Genetický kód byl popsán jako zmrazená nehoda a důvody, proč existuje pouze jedna standardní aminokyselina s přímým řetězcem (alanin), mohou být jednoduše nadbytečné s valinem, leucinem a isoleucinem. Uvádí se však, že aminokyseliny s přímým řetězcem vytvářejí mnohem stabilnější alfa helixy.

Glycin (postranní řetězec vodíku)
Alanin (methylový postranní řetězec)

Homoalanin nebo kyselina α-aminomáselná (boční postranní řetězec)

Norvalin ( n -propylový postranní řetězec)

Norleucin ( n -butylový postranní řetězec)

Homonorleucine ( n -Pentylový postranní řetězec) (ukázána kyselina heptanová)




Chalkogen
Serin, homoserin , O-methyl-homoserin a O-ethyl-homoserin mají postranní řetězec hydroxymethyl, hydroxyethyl, O-methyl-hydroxymethyl a O-methyl-hydroxyethyl. Zatímco cystein, homocystein , methionin a ethionin mají thiolové ekvivalenty. Ekvivalenty selenolu jsou selenocystein, selenohomocystein, selenomethionin a selenoethionin. Aminokyseliny s dalším chalkogenem se nacházejí také v přírodě: několik druhů, jako je Aspergillus fumigatus, Aspergillus terreus a Penicillium chrysogenum za nepřítomnosti síry, je schopno produkovat a začlenit do proteinu tellurocystein a telluromethionin.
Hydroxyglycin, aminokyselina s postranním řetězcem hydroxylu, je vysoce nestabilní.
Rozšířený genetický kód
Role
V buňkách, zejména autotrofních, se jako metabolické meziprodukty nachází několik neproteinogenních aminokyselin. Navzdory katalytické flexibilitě enzymů vázajících PLP je mnoho aminokyselin syntetizováno jako keto kyseliny ( např. 4-methyl-2-oxopentanoát na leucin) a aminovány v posledním kroku, čímž se udržuje počet neproteinogenních aminokyselin meziprodukty poměrně nízké.
Ornithin a citrulin se vyskytují v cyklu močoviny , což je součást katabolismu aminokyselin (viz níže).
Kromě primárního metabolismu je prekurzory nebo finální produkcí v sekundárním metabolismu několik neproteinogenních aminokyselin za vzniku malých sloučenin nebo neribozomálních peptidů (jako jsou některé toxiny ).
Posttranslačně začleněn do proteinu
Přestože nejsou genetickým kódem kódovány jako proteinogenní aminokyseliny, některé nestandardní aminokyseliny se přesto v bílkovinách nacházejí. Ty jsou tvořeny posttranslační modifikací postranních řetězců standardních aminokyselin přítomných v cílovém proteinu. Tyto modifikace jsou často zásadní pro funkci nebo regulaci proteinu; například v gama-karboxyglutamát karboxylace z glutamátu umožňuje lepší vázání kationtů vápníku , a hydroxyprolinu hydroxylace z prolinu je rozhodující pro udržení pojivové tkáně . Dalším příkladem je tvorba hypusinu ve faktoru iniciace translace EIF5A prostřednictvím modifikace lysinového zbytku. Takové modifikace mohou také určit lokalizaci proteinu, např. Přidání dlouhých hydrofobních skupin může způsobit, že se protein naváže na fosfolipidovou membránu.

Kyselina karboxyglutamová. Zatímco kyselina glutamová má jednu y-karboxylovou skupinu, karboxyglutamová kyselina má dvě.

Hydroxyprolin. Tato imino kyselina se liší od prolinu díky hydroxylové skupině na uhlíku 4.
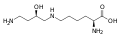
Hypusin . Tato aminokyselina se získá přidáním 4-aminobutylové skupiny k e-aminoskupině lysinu (získává se ze spermidinu )


-Pyroglutamic_acid_Structural_Formulae.png)
Existuje nějaký předběžný důkaz, že kyselina aminomalonová může být v proteinu přítomna, pravděpodobně nesprávným začleněním .
Toxické analogy
Několik neproteinogenních aminokyselin je toxických díky své schopnosti napodobovat určité vlastnosti proteinogenních aminokyselin, jako je thialysin . Některé neproteinogenní aminokyseliny jsou neurotoxické napodobováním aminokyselin používaných jako neurotransmitery (tj. Nikoli pro biosyntézu proteinů), např. Kyselina Quisqualic , canavanine nebo azetidine-2-karboxylová kyselina . Cefalosporin C má hlavní řetězec kyseliny a-aminoadipové (homoglutamát), který je amidován cefalosporinovou skupinou. Penicilamin je terapeutická aminokyselina, jejíž způsob účinku není znám.

Thialysin

Kyselina quisqualic

Canavanine

kyselina azetidin-2-karboxylová

Cefalosporin C.

Penicilamin

-Azetidine-2-carboxylate.png)

Přirozeně se vyskytující kyanotoxiny mohou také zahrnovat neproteinogenní aminokyseliny. Mikrocystin a nodularin jsou například odvozeny od ADDA , beta-aminokyseliny.
Ne aminokyseliny
Taurin je aminosulfonová kyselina a ne aminokarboxylová kyselina, ale někdy se za ni považuje, protože množství potřebná k potlačení auxotrofu v určitých organismech (např. Kočkách) jsou bližší množstvím „esenciálních aminokyselin“ (aminokyselinová auxotrofie) než vitamíny (kofaktorová auxotrofie).
Osmolyty, sarkosin a glycin betain jsou odvozeny z aminokyselin, ale mají sekundární a kvartérní amin.