
Sponge spicule - Sponge spicule

.jpg)
Spikuly jsou strukturální prvky, které se nacházejí ve většině hub . Piškoty jsou vyrobeny z uhličitanu vápenatého nebo oxidu křemičitého . Velké spikuly viditelné pouhým okem se označují jako megaskléry , zatímco menší, mikroskopické se nazývají mikroskléry . Síťování mnoha spikul slouží jako kostra houby, a tak poskytuje strukturální podporu a potenciálně obranu před predátory. Složení, velikost a tvar spikul jsou hlavními postavami systematiky a taxonomie hub .
Přehled
| Část série související s |
| Biomineralizace |
|---|
 |
Houby tvoří kmen zvaný Porifera (z latinského porus znamená „pór“ a -fer znamená „ložisko“). Porifera jsou druhově bohatá kladu nejranějších (nejvíce bazálních ) zvířat. Jsou distribuovány po celém světě s různorodými ekologiemi a funkcemi a mají rekord v délce minimálně celého fanerozoika . Většina houbiček produkuje kostry tvořené spikulami, strukturálními prvky, které se vyvíjejí v nejrůznějších velikostech a trojrozměrných tvarech. Mezi čtyřmi podtypy Porifery tři ( Demospongiae , Hexactinellida a Homoscleromorpha ) produkují kostry amorfního oxidu křemičitého a jeden ( Calcarea ) z hořčíku-kalcitu. Právě tyto kostry jsou složeny z prvků zvaných spicules . Tyto morfologie z fragmentů jsou často jedinečné clade- nebo dokonce druh úrovně taxonů díky čemuž jsou obzvláště užitečné v taxonomických úkolů.
Hloubka oceánu je hojně osídlena velmi různorodou faunou a flórou, odkud expedice Challenger (1873–1876) ocenila bohatou sbírku sklivcových hub (Hexactinellida). Byly popsány Schulzem a představují fylogeneticky nejstarší třídu křemičitých hub. Jsou okouzlující díky svému výraznému tělesnému plánu, který se spoléhá na filigránskou kostru. Je konstruován řadou morfologicky určených prvků, spikul. Později, během německé hlubinné expedice „ Valdivia “ (1898-1899), mohl Schulze popsat největší křemičitou hexactinellidovou houbu na Zemi, až tři metry vysokou Monorhaphis chuni , která vyvíjí stejně největší struktury z biosiliky , obří bazální spikuly (tři metry vysoké a jeden centimetr silné). S takovými spikulami jako modelem by mohly být zpracovány základní znalosti o morfologii, formování a vývoji kosterních prvků. Spikuly jsou tvořeny proteinovým lešením, které zprostředkovává tvorbu křemičitých lamel, ve kterých jsou proteiny obaleny. Kolem axiálního kanálu lze soustředně uspořádat až osm set 5 až 10 μm silných lamel. Oxid křemičitý je složen z téměř čistého křemíku a kyslíku a poskytuje mu neobvyklé optofyzické vlastnosti, které jsou lepší než vlastnosti umělých vlnovodů .
V posledním desetiletí, jako fylogeneticky nejstarší živočišný kmen, se Porifera (houby) těší zvláštnímu zájmu. Především kvůli zavedení molekulárně biologických technik byly vypracovány solidní důkazy, které naznačovaly, že tento kmen obsahuje hojnost nových informací pro pochopení dynamiky evolučních procesů, ke kterým došlo během ediacaranu , doby před kambrijskou explozí, která může být se datuje přibližně před 540 miliony let. Kromě toho jsou druhy tohoto kmene bohatými a užitečnými zdroji pro bioprospecting .
Houby jsou nejjednodušší mnohobuněčná zvířata, která jsou podle Granta seskupena do kmene Porifera. Grant popsal tyto přisedlé, mořské živočichy, které mají být postaveny jen z měkkého, houbovitého (amorfně tvarovaného) materiálu. Později, s objevením skleněných houbiček (třída Hexactinellida), se tento pohled drasticky změnil; pak byli považováni za „nejsilněji individualizované, radiálně symetrické“ entity. Od svého objevu byly hexactinellidy hodnoceny jako „nejcharakterističtější obyvatelé velkých hlubin, kteří soupeří“ s druhou třídou Porifery , demosponge , jejich krásou . Jejich tenká síť živých tkání je podpořena charakteristickou kostrou, jemným lešením křemičitých spikul, z nichž některé mohou být sloučeny sekundárním ukládáním oxidu křemičitého za vzniku pevného rámce. Hexactinellida spolu s Demospongiae tvoří společnou taxonomickou jednotku obsahující křemičité houby. Jejich kostry jsou postaveny z oxidu křemičitého, který je uložen ve formě amorfního opálu (SiO 2 · nH 2 O), a konstruuje řadu odlišných struktur nazývaných spikuly. Podle molekulárních údajů z genů houby, které kódují receptory a signální transdukční molekuly, byla Hexactinellida stanovena jako fylogeneticky nejstarší třída Porifera. Na základě zjištění, že Porifera sdílí jednoho společného předka, Urmetazoa , s ostatními zvířaty, bylo vyvozeno, že tato zvířata představují nejstarší, dosud existující zvířecí taxon. A co víc, vznik těchto zvířat lze vypočítat před 650–665 miliony let [Ma], což je datum, které bylo potvrzeno záznamy o zkamenělinách. Porifera tedy musela žít již před hranicí Ediacaran-Cambrian , 542 Ma, a proto jejich objasněná genetická sada nástrojů může přispět k porozumění také ediakarské měkké tělesné biotě , jak ji načrtl Pilcher. Právě evoluční novinka, tvorba tvrdé kostry, významně přispěla k ozáření zvířat v pozdním proterozoiku a konstrukci metazoanského tělesného plánu. Později v evoluci po období Ediacaran 10.1098/rspb.1999.0605 se objevila třetí třída Porifery, Calcarea, která obsahuje kostru uhličitanu vápenatého.
Houby hexactinellid se vyznačují křemičitými spikulami, které vykazují hexaktinickou, triaxonickou (kubickou) symetrii nebo morfologii odvozenou redukcí ze základních stavebních plánů spicul. Jejich tvary těla jsou méně variabilní a strukturovanější než u Demospongiae. Hexactinellida byly rozděleny do dvou hlavních linií, Amphidiscophora a Hexasterophora. Mají tvar trychtýře až šálku a dosahují stability svých těl pomocí pinulárních pentaktinů a zřídka hexaktinů, zatímco fixaci k substrátu udržují bazálie (monaktiny). Je to variace v basalia, která dává Amphidiscophora jejich výraznou morfologii. Basalia mohou být zabaleny nebo dokonce baleny dohromady. Nejvýznamnějšími druhy tohoto řádu jsou Monorhaphis a Hyalonema kvůli jejich velikostem. Druhý řád hexaktinellidů představuje Hexasterophora, které obsahují tuhou diktyonální strukturu pocházející z jednoduchých hexaktinů. Jejich tělesné plány mají obvykle rozvětvené a anastomózní formy s koncovými oskulárními ploténkami. Nejznámějším příkladem je Euplectella aspergillum.
Typy spicule

Piškoty mohou být vápenaté , křemičité . Silikonové spikuly jsou někdy vloženy do sponginu .
Spikuly se nacházejí v řadě typů symetrií.
Monaxony tvoří jednoduché válce se špičatými konci. Konce přímočarých monaxonů jsou podobné, zatímco monaktinální monaxony mají různé konce: jeden špičatý, jeden zaoblený. Diaktinální monaxony jsou klasifikovány podle povahy jejich konců: oxea mají špičaté konce a strongyly jsou zaoblené. Páteřní oxea a strongyly se nazývají acanthoxea a acanthostrongyles. Monaktické monaxony mají vždy jeden špičatý konec; nazývají se styly, pokud je druhý konec tupý, tylostyly, pokud jejich tupý konec tvoří knoflík; a acanthostyly, pokud jsou pokryty ostny.
Triaxony mají tři osy; v trojicích nese každá osa podobný paprsek; u pentaktů má triaxon pět paprsků, z nichž čtyři leží v jedné rovině; a pinnules jsou pentakty s velkými trny na nerovinném paprsku.
Tetraxony mají čtyři osy a polyaxony více (popis typů, ze kterých mají být začleněny). Sigma-C spikule mají tvar C .
Dendroclones může být jedinečné zaniklé houby a jsou větvení spicules, které mohou trvat nepravidelné formy, nebo mohou tvořit struktury s I , Y nebo X tvaru.
- Megaskléry jsou velké spikuly měřící od 60 do 2 000 μm a často fungují jako hlavní nosné prvky v kostře.
- Acanthostyles jsou ostnaté styly.
- Anatriaeny, orthotriaeny a protriaeny jsou triaeny - megaskléry s jedním dlouhým a třemi krátkými paprsky.
- Strongyly jsou megascléry s oběma konci tupými nebo zaoblenými.
- Styly jsou megascleres s jedním koncem špičatým a druhým koncem zaobleným.
- Tornotes jsou megascleres s konci ve tvaru kopí.
- Tyloty jsou megaskléry s knoflíky na obou koncích.
- Mikrosklery jsou malé spikuly o velikosti 10–60 μm a jsou roztroušeny po tkáni a nejsou součástí hlavního nosného prvku.
- Chelae jsou mikroskleny s lopatkovitými strukturami na koncích. Anisochely jsou mikroskleny s rozdílnými konci. Isochelas jsou mikroskleny se dvěma podobnými konci.
- Euasters jsou hvězdicovité mikroskleny s více paprsky vyzařujícími ze společného centra. Příklady jsou oxyasters (euasters se špičatými paprsky) nebo sterrasters (kuličkovité euasters).
- Kleště jsou mikroskleny ohnuté zpět na sebe.
- Microstrongyles jsou malé pruty s oběma konci tupými nebo zaoblenými.
- Microxeas jsou malé tyče s oběma konci špičatými.
- Sigmas jsou mikroskleny ve tvaru „C“ nebo „S“.
Formace
Spikuly jsou tvořeny sklerocyty , které jsou odvozeny z archaocytů . Sklerocyt začíná organickým vláknem a přidává do něj oxid křemičitý. Spikuly se obecně prodlužují rychlostí 1 až 10 μm za hodinu. Jakmile spikula dosáhne určité délky, vyčnívá z těla buněk sklerocytů , ale zůstává uvnitř buněčné membrány . Příležitostně mohou sklerocyty začít s druhým spiculeem, zatímco první stále probíhá.
Vápenaté spikuly



Biomineralizace zvířat je kontrolovaný proces a vede k výrobě minerálně -organických kompozitních materiálů, které se tvarově a materiálovými vlastnostmi výrazně liší od čistě anorganických protějšků. Schopnost vytvářet funkční biominerály, jako jsou endoskeletony a exoskeletony , ochranné skořápky nebo zuby, byla významným krokem v evoluci zvířat. Biomineralizace uhličitanu vápenatého, nejrozšířenější typ mezi živočišnými kmeny, se vyvinula několikrát nezávisle, což vedlo k vícenásobnému náboru stejných genů pro biomineralizaci v různých liniích.
Mezi těmito geny jsou členové rodiny genů rodiny alfa -karboanhydrázy (CA) nezbytní pro biomineralizaci. CA jsou enzymy vázající zinek, které katalyzují reverzibilní přeměnu oxidu uhličitého a vody na hydrogenuhličitan a jeden proton. Vazba na zinek je zprostředkována třemi histidinovými zbytky nezbytnými pro katalytickou funkci proteinu . CA se podílejí na mnoha fyziologických procesech vyžadujících regulaci iontů nebo transport uhlíku, přičemž oba jsou zásadní pro řízené srážení biominerálů uhličitanu. U savců, kde jsou nejlépe studováni, je 16 různých CA exprimováno ve specifických tkáních a aktivní v definovaných subcelulárních kompartmentech. Lze rozlišit cytosolické , mitochondriální , membránové a sekretované formy CA a tyto skupiny se v různých zvířecích skupinách rozšířily a redukovaly. Specifické CA se podílejí na biomineralizaci uhličitanu v odlišných liniích zvířat, včetně hub.
Mezi existujícími houbami mohou pouze vápenaté houby produkovat kalcitové spicules, zatímco spicules jiných tříd jsou křemičité. Některé linie mezi demosponge a několika calcareany mají masivní bazální kostry uhličitanu vápenatého, takzvané koralínové houby nebo sklerosponge . Tyto biomineralizing CA používají uhličitan produkující demosponges nejsou ortologní do CAS podílejí na tvorbě spicule vápnitých houby, což naznačuje, že tyto dva typy biomineralizace vyvíjely nezávisle. Toto pozorování souhlasí s myšlenkou, že tvorba kalcitických spicul je evoluční inovací vápenatých houbiček.
Tvary vápnitých houbových spikul jsou jednoduché ve srovnání s někdy velmi komplikovanými křemičitými spikulami, které se vyskytují v jiných třídách houbiček. Až na několik výjimek mohou být vápenaté houbové spikuly tří základních typů: monaxonické, dvouhroté diaktiny, triaktiny se třemi paprsky spikuly a čtyřpaprskové tetraktiny. Specializované buňky, sklerocyty, produkují tyto spikuly a pouze několik sklerocytů interaguje za vzniku jednoho specifického spikula: dva sklerocyty produkují diaktin, šest sklerocytů tvoří triaktin a sedm tetraktinů. Dvojice sklerocytů se podílí na růstu každého aktinu těchto spikul. Po počáteční fázi takzvaná zakladatelská buňka podporuje prodloužení aktinu, druhá, takzvaná zahušťovací buňka u některých, ale ne u všech druhů, ukládá na aktin další uhličitan vápenatý, protože migruje zpět k zakladatelské buňce. Vápenaté houby mohou mít ve svém těle pouze jeden nebo jakoukoli kombinaci tří typů spicule a v mnoha případech jsou určité typy spicule omezeny na konkrétní části těla. To naznačuje, že tvorba spicule je pod přísnou genetickou kontrolou u vápenatých houbiček a konkrétní CA hrají zásadní roli v této genetické kontrole
Silikátové spikuly

Geodia atlantica ; h) je hilum; šipka ukazuje na hladkou růžici tvořenou paprsky. Levá stupnice 20 μm, pravá stupnice 2 μm.

Geodia macandrewii , levá stupnice 50 μm, pravá stupnice 2 μm
Silikátové spikuly v demospongech existují v různých tvarech, z nichž některé vypadají jako drobné skleněné koule. Říká se jim sterrasters, pokud patří do čeledi Geodiidae, a selenasters, pokud patří do čeledi Placospongiidae .
Silikátové spikuly poprvé popsal a ilustroval v roce 1753 Vitaliano Donati , který je našel v druhu Geodia cydonium z Jaderského moře : tyto spikuly nazval „malé kuličky“. Od houbových taxonomů se jim později říká globulární krystaloidy, globulní spikuly nebo globostelláty, až do roku 1888, kdy William Sollas konečně razil termín „sterraster“ z řeckých sterros, což znamená „pevný“ nebo „pevný“ - viz diagram vpravo. Mezitím jsou podobné koule ve tvaru koule pozorovány u jiného rodu, Placospongia , a ty jsou zpočátku považovány za „sterrastery“, než Richard Hanitsch v roce 1895 razí termín „selenaster“ pro tyto různé spikuly (pocházející z řeckého selenu pro „měsíc“) , odkazující na tvar „půlměsíce“). Nakonec von Lendenfeld v roce 1910 vytvořil další termín „aspidaster“, přesvědčený, že zploštělé sterrastery v rodu Erylus se výrazně liší od těch v Geodii .
Geodiidae dnes představují velmi různorodou rodinu houby s více než 340 druhy, vyskytující se v mělkých až hlubokých vodách po celém světě kromě Antarktidy. Sterrastery/aspidaster spicules jsou v současné době hlavní synapomorfií Geodiidae. Rodina v současné době zahrnuje pět rodů se sterrastery a několik dalších, které sekundárně ztratily sterrastery. Geodia může být masivní zvířata více než jeden metr v průměru.
.jpg)
Znázorněno vlevo: Největší strukturou biosiliky na Zemi je obří bazální spiku z hlubinné skleněné houby Monorhaphis chuni .
Selenasters jsou hlavní synapomorphy of Placospongia (rodinné Placospongiidae , objednat Clionaida ), což je dobře podporován monophyletic rodu z mělkých mírných / tropických vodách po celém světě. Nejde o příliš různorodý rod, v současnosti je popsáno pouze 10 druhů (WPD) a hrstka nepopsaných druhů . Druhy placospongia jsou obvykle malé, zakrývající se a nikdy se nevyskytují ve vysokých hustotách.
Sterrastery/selenasters jsou dostatečně velké na to, aby mohly podrobně prozkoumat jejich povrchy optickým mikroskopem. Použití rastrovacího elektronového mikroskopu (SEM) však umožnilo výrazně lepší porozumění mikroornamentacím povrchu. Objevilo se také několik popisných termínů, které podrobněji popisují a porovnávají mikroornamentace těchto kulovitých spikul. polyaxiální spikuly, jako jsou sterrastery a aspidasters, jsou výsledkem srostlých „aktinů“ (= větví asterů, z řečtiny „hvězda“), později pokrytých „rozetami“ z různých „paprsků“. „ Hilum “ (latinsky „maličkost“ nebo „maličkost“ nebo „oko fazole“) je malá oblast bez rozet nebo jakéhokoli druhu povrchového vzoru. Neexistují žádné konkrétní termíny popisující povrch selenasterů, kromě „hilum“, který je také přítomen. Ačkoli se zdá, že mezi druhy nejsou významné rozdíly ve velikosti růžic a hilu, všimli jsme si, že růžice mohou být hladké nebo bradavičnaté, a předpokládali, že tento charakter může mít fylogenetickou hodnotu, pokud bude studován v širším měřítku. Kromě toho se zdálo, že morfologie růžice je proměnlivá mezi Geodia , Pachymatisma a Caminella, což naznačuje, že podrobnější studie povrchu sterraster/aspidaster by potenciálně přinesla nové znaky pro identifikaci rodů Geodiidae.
Největší strukturou biosiliky na Zemi je obří bazální pupínek hlubinné skleněné houby Monorhaphis chuni . Schéma vpravo ukazuje:
- (a) Mladé exempláře M. chuni ukotvené k bahnitému substrátu jediným obrovským bazálním hrotem (gbs). Tělo (bo) obklopuje spicule jako souvislý kulatý válec.
- (b) Růstové fáze přisedlého zvířete s jeho GBS (gbs), které jej ukotví k substrátu a drží okolní měkké tělo (bo). Charakteristický habitus zobrazuje lineárně uspořádané velké síňové otvory (at) o průměru přibližně 2 cm. S růstem měkké tělo v bazální oblasti odumírá a odhaluje holý GBS (a až c).
- c) Část těla (bo) s jeho síňovými otvory (at). Povrch těla je proložen přijímacími otvory, které umožňují nepřetržitý průtok vody vnitřními kanály, které ústí do oskul, které jsou centralizovány v síňových otvorech, sítových deskách.
- d) M. chuni ve svém přirozeném prostředí s měkkým dnem bathyal svahů u Nové Kaledonie. Exempláři žijí v hloubce 800–1 000 m [23]. V této oblasti se houba vyskytuje při hustotě osídlení 1–2 jedinců na m 2 . Zvířata dosahují velikosti kolem 1 m na délku.
- e) Kresby různých skleněných houbiček ( hexactinellids ).
Spikule „životní cyklus“

Od formace po depozici
Tvorba spikul je řízena geneticky. Ve většině případů je první růstová fáze intracelulární; začíná ve sklerocytech (améboidních buňkách zodpovědných za tvorbu spikuly) v mesohylu a je zprostředkován silikátem, speciálním enzymem, který iniciuje tvorbu axiálního vlákna (uchyceného v axiálním kanálu), které poskytuje svislou osu spikuly. Axiální kanál je naplněn organickým proteinovým materiálem, který se obvykle rozprostírá ke špičce nově vytvořeného spikula. Průřez axiálního kanálu se liší napříč hlavními houbovými plátky, které produkují křemičité spikuly (u demospongů je trojúhelníkový, u homoskleromorfů nepravidelný a u hexactinellidů čtyřúhelníkový. U calcareanů (produkujících vápnité spikuly) není axiální kanál vyvinut. Geometrie a délka axiálního vlákna určuje tvar spikuly. V desmoidních spikulech „lithistidů“ (neformální skupina demospongů s kloubovými kostrami) je však axiální filament kratší než ramena spicule a je možné, že se jedná pouze o organické molekuly zapojený do procesu tvorby spicule.
Během tvorby křemičitých spikul (Calcarea zobrazuje různé mechanismy biomineralizace spicule) houby získávají křemík ve formě rozpustné kyseliny křemičité a ukládají jej kolem osového vlákna ve speciální membráně zvané silicalemma. Oxid křemičitý je nejprve rozložen jako malé 2 μm granule, které jsou fúzovány do větších sfér (nebo fúzovány dohromady v rámci procesu biologického spékání v Hexactinellida. Po nějaké době se přidá amorfní oxid křemičitý, čímž se vytvoří rovnoměrně uložené koncentrické vrstvy, oddělené od sebe ultratenkými organickými látkami V této fázi jsou nezralé spikuly vylučovány ze sklerocytů a pokryty pseudopodií jedné až několika buněk a pokračuje proces ukládání oxidu křemičitého a růstu spikuly.
Po dokončení depozice oxidu křemičitého (nebo během této fáze) je spikula transportována na správné místo v těle houby procházením mezohylových buněk, kde spongocyty kolem sebe vylučují sponginové fibrily a spojují je se sousedními spikulami. U některých hexactinellidů, které se vyznačují tuhou kostrou, se zdá, že fúze spikul probíhá paralelně se sekrecí spicule.
Když jsou houby živé, jejich spikuly poskytují strukturální „rámec“. Po jejich smrti se tělo a struktura skeletu, zejména u demospongů, ve kterých jsou spikuly navzájem spojeny pouze rychle se kazícími kolagenovými vlákny, rychle rozpadají a nechávají spicules „volné“. Z tohoto důvodu jsou houby ve fosilních záznamech zřídka zcela zachovány. Jejich spikuly jsou však začleněny do sedimentů a často se stávají jednou z hlavních složek sedimentárních hornin. Někdy se spikuly hromadí v obrovských aglomeracích, kterým se říká rohože nebo postele. Tyto akumulace jsou charakteristické pro polární vody. Spikuly mohou zkamenět za vzniku zvláštního druhu hornin nazývaných spiculity („ spongillity “ pro sladkovodní houby); tyto druhy hornin jsou známé po celém světě a byly vytvořeny během celého fanerozoika. Biosiliceous sedimentace občas vede k tvorbě spiculitických rohovců (v takzvaných skleněných rampách), které jsou zaznamenány od permu do eocénu v mnoha částech světa.
Spiculity
Když se mrtvá těla houby rozpadají, spikuly se začlení do mořských sedimentů a někdy se hromadí v obrovských aglomeracích, které se nazývají rohože nebo lůžka, nebo zkamenějí a vytvářejí speciální typ hornin nazývaný spiculity.
Záznamy o fosilních a subfosilních houbových spikulích jsou mimořádně bohaté a často slouží jako základ pro dalekosáhlé rekonstrukce piškotových komunit, ačkoli spikule jsou také nositeli významných ekologických a environmentálních informací. Specifické požadavky a preference hub lze použít k interpretaci prostředí, ve kterém žili, a k rekonstrukci oscilací v hloubkách vody, pH, teplot a dalších parametrů, které poskytují snímky minulých klimatických podmínek. Na druhé straně jsou kompozice izotopů křemíku v spikulech (δ30Si) stále častěji používány k odhadu hladiny kyseliny křemičité v mořském prostředí v celé geologické historii, což umožňuje rekonstrukci minulého cyklu oxidu křemičitého a cirkulace oceánu .
Spikuly poskytují strukturální podporu pro udržení svislé polohy těla, minimalizují metabolické náklady na výměnu vody a mohou dokonce odradit predátory. Často se vyvíjejí v různých velikostech a široké škále trojrozměrných tvarů, přičemž mnohé z nich jsou jedinečné pro taxony na kladu nebo dokonce na úrovni druhů. Demosponge jsou charakterizovány spikulami monaxonické nebo tetraxonické symetrie. Hexactinellids produkují spikuly hexaktinické nebo triaxonické (kubické) symetrie nebo tvary, které jsou z takových morpologií jasně odvozeny. Jehliček homoscleromorphs představují podivné tetractines ( calthrops ) a jejich deriváty, které vznikají prostřednictvím snížení nebo rozvětvení z clads. Spicules of Calcarea se vyrábějí ve třech základních formách: diaktiny, triaktiny a tetraktiny.
Minerální složení houbových spikul činí z těchto struktur nejodolnější části těl houbiček a zajišťuje schopnost spikul odolávat různým tafonomickým procesům , což má za následek, že často představují jediný důkaz přítomnosti některých houb v ekosystému. I když jsou houby často známé z bohatých asambláží tělesně zachovaných exemplářů, významnou část jejich fosilních a subfosilních záznamů představují také jejich spikuly. S ohledem na to mohou mít spikule zásadní význam pro rekonstrukce zaniklých nebo kryptických (skrývajících se v děložních čípcích a jeskyních) komunit houby; a skutečně byly zkoumány zejména s ohledem na jejich taxonomický význam. Morfologie spikul a jejich uspořádání spolu s dalšími důležitými vlastnostmi houby, jako je tvar, konzistence a barva, jsou zásadní při identifikaci houbiček.
Na rozdíl od fosilií houby s plným tělem jsou spikuly běžné v mnoha depozičních prostředích . Jejich význam je však často podceňován, což je většinou způsobeno obtížemi při přiřazování disociovaných spicul taxonům houby nebo kvůli nedostatku materiálu.
Interakce se světlem
Výzkum na Euplectella aspergillum (Venušin květinový koš) prokázal, že spikuly některých hlubinných hub mají podobné rysy jako optické vlákno . Kromě schopnosti zachytit a transportovat světlo mají tyto spikuly řadu výhod oproti komerčním drátům z optických vláken. Jsou silnější, snáze odolávají stresu a vytvářejí si vlastní podpůrné prvky. Nízkoteplotní tvorba spikul ve srovnání s vysokoteplotním procesem natahování komerčních optických vláken umožňuje přidání nečistot, které zlepšují index lomu . Tyto spikuly mají navíc na koncích zabudované čočky, které shromažďují a zaostřují světlo v tmavých podmínkách. Bylo teoretizováno, že tato schopnost může fungovat jako světelný zdroj pro symbiotické řasy (jako u Rosella racovitzae ) nebo jako atraktor pro krevety, které žijí ve Venušině květovém koši. Nebylo však přijato konečné rozhodnutí; může se stát, že světelné schopnosti jsou pouze náhodnou vlastností čistě strukturálního prvku. Spicules trychtýř světlo hluboko uvnitř mořské houby.
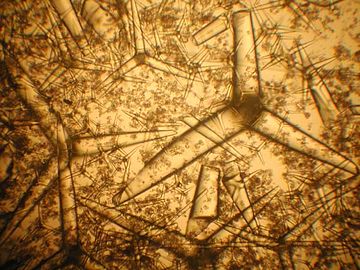
Spongelové spikuly

Spicules of sponge ( SEM )

Síť houbových spikul

.JPG)

Viz také
Reference
Další reference
- Shoham, Erez; Prohaska, Thomas; Barkay, Zahava; Zitek, Andreas; Benayahu, Yehuda (2019). „Měkké korály tvoří aragonitem vysrážený sloupcový spiculit v mezofotických útesech“ . Vědecké zprávy . 9 (1): 1241. Bibcode : 2019NatSR ... 9.1241S . doi : 10,1038/s41598-018-37696-z . PMC 6362064 . PMID 30718658 .